条文本

摘要
客观的脂质代谢异常是结直肠癌(CRC)的一个标志。角鲨烯环氧化酶(SQLE)是胆固醇生物合成中的限速酶,在CRC中表达上调。在这里,我们的目的是确定SQLE的致癌功能及其与肠道菌群在促进结直肠肿瘤发生中的相互作用。
设计对来自两个队列的相邻正常组织和CRC进行分析(n=202)。通过将Rosa26-lsl-Sqle小鼠与Cdx2-Cre小鼠杂交,生成结肠特异性Sqle转基因(Sqle tg)小鼠。收集粪便进行宏基因组学和代谢组学分析。
结果结直肠癌中SQLE信使RNA和蛋白表达上调(p<0.01),预示结直肠癌患者生存期较差。SQLE通过诱导细胞周期进程和抑制细胞凋亡促进CRC细胞增殖。在氮氧甲烷诱导的CRC模型中,Sqle tg小鼠与野生型小鼠相比,肿瘤发生明显增加(p<0.01)。综合宏基因组学和代谢组学分析揭示了富致病菌的Sqle tg小鼠的肠道失调,这与次生胆汁酸的增加有关。与次生胆汁酸的有害作用一致,Sqle tg小鼠的肠道屏障功能受损,紧密连接蛋白Jam-c和occludin减少。与对照组小鼠粪便相比,Sqle tg小鼠粪便移植无菌小鼠的肠道屏障功能受损,并刺激细胞增殖。最后,我们证明,通过与奥沙利铂和5-氟尿嘧啶协同作用,特比萘芬(一种SQLE抑制剂)可用于CRC,以抑制CRC生长。
结论本研究表明,SQLE通过细胞内作用和调节肠道微生物-代谢产物轴介导肿瘤发生。SQLE是结直肠癌的治疗靶点和预后标志物。
- 结肠直肠癌
- 结直肠肿瘤
- 结肠微生物区系
数据可用性声明
根据合理的要求提供数据。所有资料均可向通讯作者索取。
这是一篇开放获取的文章,按照创作共用署名非商业性(CC BY-NC 4.0)许可发布,该许可允许其他人以非商业性的方式发布、混编、改编、构建本作品,并以不同的条款授权他们的衍生作品,前提是原创作品被正确引用,给予适当的荣誉,任何更改都被注明,且使用是非商业性的。看到的:http://creativecommons.org/licenses/by-nc/4.0/.
来自Altmetric.com的统计
本研究的意义
关于这个问题,我们已经知道了什么?
脂质代谢异常是结直肠癌(CRC)的一个标志。
在结直肠癌中,胆固醇代谢经常上调,但其功能及其与肠道微生物群的潜在相互作用尚不清楚。
新的发现是什么?
角鲨烯环氧化酶(SQLE)在结直肠癌中过表达,其高表达预示着较差的生存率。
结肠靶向Sqle在小鼠中转基因表达促进偶氮甲烷诱导结直肠癌的成瘤。
综合宏基因组学和代谢组学分析表明结肠特异性Sqle转基因小鼠体内微生物群失调,致病菌丰富,与次生胆汁酸产生增加有关。
紧连接蛋白Jam-c和occludin被抑制后,Sqle转基因小鼠的肠道屏障功能被破坏。
将转基因Sqle小鼠粪便移植到无菌小鼠体内可促进结肠细胞增殖和肠道屏障破坏。
SQLE抑制剂特比萘芬与化疗协同抑制结直肠癌体外和体内生长。
在可预见的未来,它会对临床实践产生怎样的影响?
以sqle -肠道微生物组轴为靶点是预防和治疗结直肠癌的潜在策略。
简介
结直肠癌(CRC)是全球第三大常见癌症,在癌症死亡率方面排名第二。儿童CRC的发病率和死亡率在年轻人中仍在增加,特别是在低收入和中等收入国家。晚期结直肠癌转移患者预后较差,耐药严重影响化疗疗效。因此,识别CRC发病机制中涉及的新基因以开发有效的治疗方法是很重要的,无论是单独治疗还是联合化疗。
脂质代谢异常越来越被认为是包括结直肠癌在内的许多癌症的标志特征。肥胖和血脂都是结直肠癌的危险因素。1 2流行病学研究还表明,高脂肪摄入会增加结直肠癌的风险。3.脂质积累通过增加活性氧和MAPK信号传导促进CRC。4脂肪酸,5个6胆固醇7 8和胆汁酸9与CRC的发病机制有关。然而,涉及CRC的脂质代谢依赖分子机制仍然知之甚少。
角鲨烯环氧化酶(SQLE)是胆固醇生物合成中的限速酶。最近的研究表明,SQLE在CRC中过度表达8 10 11;然而,SQLE在CRC发生和进展中的潜在作用仍有争议。提示SQLE是通过促进细胞生长而致癌的因子。10而6月等8显示SQLE的降低可加速CRC转移。在这项研究中,我们证明了在多个CRC队列中,与相邻正常组织相比,SQLE在CRC中表达上调,并预测患者生存期较差。
为了全面解读SQLE在结直肠癌中的作用,我们首先建立了结肠特异性SQLE转基因(tg)小鼠和SQLE敲除(KO)小鼠,并在体内验证了SQLE作为结肠癌致癌基因在结直肠癌中的作用。我们发现结肠特异性SQLE过表达促进肠道失调和代谢改变,从而导致肠道屏障受损和诱导结肠细胞增殖。最后,我们展示了特比萘芬,一种食品和药物管理局(FDA)批准的SQLE抑制剂,与5-氟尿嘧啶(5-FU)或奥沙利铂协同抑制CRC细胞的体外和体内生长。这些发现提示SQLE是CRC的一种靶向致癌因子。
材料和方法
人类CRC样本
组织学证实结直肠癌的患者样本(香港队列)在北京大学肿瘤医院接受手术时收集(n=144)。肿瘤和邻近的正常组织立即在液氮中快速冷冻。病人的特点总结在在线补充表S1.另一项组织微阵列(TMA)队列由207名在香港威尔斯亲王医院接受手术的CRC患者组成。病人的特点总结在在线补充表S2.使用一个公开的队列(GSE17538)进行验证,包括232例CRC患者(在线补充表S3).癌症基因组图谱(TCGA)队列数据集通过UCSC Xena浏览器访问(https://xenabrowser.net/),使用来自rna测序的50对相邻正常和肿瘤样本。GDS4382由17个配对的相邻正常和肿瘤样本组成,通过Affymetrix人类基因组U133 Plus 2.0阵列(https://www.ncbi.nlm.nih.gov/sites/GDSbrowser?acc=GDS4382).
结肠特异性Sqle转基因小鼠模型
Sqle tg小鼠(rosa26 - pcap -loxp-stop-loxp- Sqle)是我们之前建立的。12为了驱动SQLE的结肠特异性表达,将SQLE tg小鼠与Cdx2-Cre交叉ERT2老鼠。为了激活SQLE过表达,先给6周龄SQLE tg小鼠腹腔注射他莫昔芬(100 mg/kg,腹腔注射),每日4次,以激活CreERT2.为了诱导Sqle tg小鼠发生结直肠癌,我们采用了化学诱导的CRC模型。简单地说,偶氮甲烷(AOM, 10 mg/kg,腹腔注射)给小鼠6次,每周1次。第一次注射AOM后第24周处死小鼠。记录肿瘤的数量和大小。所有组织立即保存在10%福尔马林或−80°C冰箱中。组织学评分后,H&E染色的病理学家盲目的样品的性质。
Sqle敲除小鼠模型
采用CRISPR/Cas9技术制备全体Sqle KO小鼠。杂合子Sqle KO小鼠(Sqle+ /−由于纯合子的KO小鼠无法存活,因此采用aom -葡聚糖硫酸钠(DSS)模型。为了诱导CRC, Sqle+ /−野生型小鼠在6周龄时腹腔注射AOM 1次(10 mg/kg)。AOM注射1周后,进行3个循环DSS处理(在饮用水中加入2%,持续5天),每个循环之间间隔2周。第一次注射AOM后80天处死小鼠。测量肿瘤的数量和大小。所有组织立即保存在10%福尔马林或−80°C冰箱中。
异种移植模型
SW1116和LOVO细胞表达空载体或SQLE质粒(1×107在4周龄Balb/c裸鼠左背皮下注射0.1 mL磷酸缓冲盐水(PBS)细胞。用数字卡尺测量皮下肿瘤大小。4周后处死小鼠。HT29细胞(1×1070.1 mL PBS)皮下注射于裸鼠背侧。小鼠被随机分组并接受:(1)灌胃;特比萘芬(80mg /kg,口服,每日);(3) 5-FU (30mg /kg,腹腔注射,2次/周);(4)奥沙利铂(10 mg/kg,腹腔注射,每周1次);(5)特比萘芬+5- fu和(6)特比萘芬+奥沙利铂。观察肿瘤大小,4周后处死小鼠。
体内肠通透性试验
为测定小鼠体内肠通透性,小鼠饥饿过夜,灌胃异硫氰酸荧光素(FITC)-葡聚糖(Sigma) (44 mg/100 g体重)。4 h后麻醉小鼠,心脏穿刺采血,处死小鼠。将血清与全血分离,稀释后用荧光分光光度法(BioTek)测定FITC荧光,激发和发射分别为485 nm和528 nm。以连续稀释的fitc -葡聚糖为标准。
强迫metagenome测序
采集Sqle tg小鼠和野生型凋落物粪便,采用PowerFecal DNA分离试剂盒提取粪便。使用Nextera XT适配器生成扩增文库,使用HiSeq Illuminex 2500进行测序,获得每个样品深度为10G碱基对的DNA (125 bp)配对端reads。在本研究中,所有粪便宏基因组都使用了相同的注释管道。为了进行数据分析,使用BWA和SAM工具进行宿主DNA去除和reads质量过滤。13日14在此之后,Kraken2和Bracken使用高质量的读取来获得物种级别的分类学分析。15日16用R包DESeq2进行种级细菌的差异丰度分析。17调整后p<0.05和log2(倍变)>1的菌种被认为有统计学意义。主坐标分析采用R包phyloseq和vegan。18
统计分析
所有统计检验均采用SPSS(19.0版)或GraphPad软件进行。数据以平均值±标准差表示。细胞活力测定和肿瘤生长速率评价采用重复测量方差分析。采用配对双尾Student 's t检验分析临床特征对SQLE信使RNA (mRNA)表达的影响及两组间比较。应用Pearson χ分析临床特征对SQLE蛋白表达的影响2或者费雪精确判别法。为了评估SQLE与预后之间的相关性,总生存期(OS)被定义为从手术之日到死亡之日的时间。利用Kaplan-Meier分析,建立OS曲线。多因素分析采用Cox比例危险回归分析。为了确定SQLE对OS的显著性,采用log-rank检验。P值<0.05认为有统计学意义。
中提供了其他方法在线补充信息.
结果
SQLE在结直肠癌患者中过表达
我们分析了三个独立CRC队列(内部香港队列、TCGA和GDS4382)中SQLE的mRNA表达。香港队列的临床特征见在线补充表S1.定量PCR (qPCR)结果显示,与相邻正常组织相比,原发性结直肠癌中SQLE mRNA高度上调(n=141, p<0.0001)。图1一个).SQLE mRNA表达与晚期肿瘤、淋巴结、转移分期及远处转移有显著相关性(p<0.05) (在线补充表S1).其过表达在TCGA中得到验证(n=50;p<0.0001)和GDS4382队列(n=17;p < 0.001) (图1一个).在蛋白质水平,免疫组化(IHC)证实了与邻近正常组织相比,CRC肿瘤中SQLE蛋白的过表达(p<0.01) (图1 b).因此,SQLE过表达式是CRC中经常发生的事件。
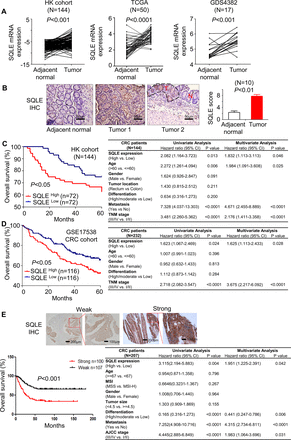
角鲨烯环氧化酶(SQLE)在结直肠癌(CRC)中上调,其表达预示着较差的生存率。(A)在配对肿瘤和邻近正常组织中测定SQLE信使RNA (mRNA)的表达,这些组织来自香港(HK)队列(北京,n=144)、癌症基因组图谱(TCGA)队列(n=50)和GDS4382队列(n=17)。配对双尾学生t检验用于计算p值。(B)用免疫组化法(IHC)检测成对肿瘤和相邻正常组织(n=10)中SQLE蛋白的表达。配对双尾学生t检验用于计算p值。比例尺,50µm。(C) Kaplan-Meier曲线分析显示,在HK队列中,高SQLE mRNA的CRC患者5年生存率明显低于低SQLE mRNA的患者(左)。考克斯回归分析(右)。(D)根据Kaplan-Meier曲线(左)和Cox回归分析(右),GSE17538队列中SQLE mRNA表达与生存率差相关。(E)组织微阵列(TMA)分析显示,高SQLE蛋白表达的CRC患者5年生存率明显较低,Kaplan-Meier曲线(左)和Cox回归分析(右)证实。 Nuclear staining of 10% was used as the cut-off value. *P<0.05; **p<0.01; ***p<0.001. AJCC, The American Joint Committee on Cancer; TNM, tumour, node, metastases.
SQLE过表达与CRC患者预后不良相关
鉴于SQLE在结直肠癌中频繁过表达,我们随后评估了SQLE在人类结直肠癌中的临床意义。在香港队列中,Kaplan-Meier曲线显示,高SQLE mRNA水平与结直肠癌患者生存率差相关(p<0.05) (图1 c).多因素Cox比例风险回归分析一致显示,高SQLE mRNA表达是CRC患者疾病特异性生存差的独立预后因素(p<0.05;人力资源1.832;95%可信区间1.113至3.113)(图1 c).我们在GSE17538队列(n=232)中验证了SQLE mRNA的预后意义(在线补充表S3)采用Kaplan-Meier曲线(p<0.05)和单因素Cox比例风险回归分析(p<0.05) (图1 d).
为了确定SQLE是否在蛋白质水平上预测患者预后,我们对207例CRC病例进行了TMA分析(图1 e).对SQLE蛋白水平的临床特征评估显示,与美国癌症联合委员会癌症分期(p<0.01)、淋巴结转移(p<0.01)和远处转移(p<0.001)显著相关(在线补充表S2),暗示SQLE可能参与了肿瘤的进展。事实上,通过Kaplan-Meier曲线分析(p<0.001), SQLE蛋白高表达与结直肠癌患者生存率差相关(p<0.001)。图1 e多变量Cox比例风险回归分析(p<0.05;人力资源1.951;95%可信区间1.225至2.391)(图1 e).我们的数据共同表明,SQLE mRNA和蛋白表达是预测CRC患者生存不良的独立预后因素。
SQLE促进CRC细胞生长
为了确定SQLE在人结直肠癌中的致癌作用,我们在结直肠癌细胞LOVO和SW1116 (图2一个),内源性SQLE水平较低(在线补充图S1).SQLE表达促进细胞生长,细胞生长曲线(p<0.01) (图2一个)和菌落形成试验(p<0.01) (图2 b).相反,我们在CRC细胞HT29和DLD1 (在线补充图S1),具有较高的内源性SQLE表达式(图2 c).SQLE短发夹RNA抑制细胞活力(p<0.01) (图2 c)和菌落形成(p<0.01) (图2 d)与shControl细胞比较。然后,在裸鼠模型上研究SQLE的致瘤作用。将稳定表达空载体或SQLE的LOVO细胞和SW1116细胞分别皮下注入裸鼠左右背侧。所示图2 e, SQLE过表达可显著增加LOVO和SW1116异种移植瘤体积(p<0.01)。SQLE耗竭也抑制了原代crc衍生类器官的增殖(图2 f).总之,SQLE促进CRC细胞在体外和体内的生长。

角鲨烯环氧化酶(SQLE)促进结直肠癌(CRC)细胞增殖。(A)在LOVO和SW1116细胞中过表达SQLE促进细胞活力和(B)集落形成。(C)在HT29和DLD1细胞中敲除SQLE显著抑制细胞活力和(D)菌落形成。(E) SQLE在体内促进肿瘤生长。SW1116和LOVO细胞(5×106转染SQLE或对照载体的细胞/肿瘤)注射给4周龄的裸鼠。与对照组相比,SQLE过表达细胞的肿瘤生长明显更快。(F) SQLE在CRC的两个主要类器官中过表达,敲除SQLE抑制了它们的生长。通过重复测量方差分析确定生长曲线之间差异的显著性。菌落形成的差异由双尾Student 's t检验决定。* P < 0.05;* * p < 0.01;* * * p < 0.001;* * * * p < 0.0001。所有实验均进行3次,每组3个重复。
SQLE抑制CRC细胞凋亡,促进细胞周期进程
接下来,我们测定了SQLE在CRC细胞系中的细胞动力学效应。用annexin V/7-氨基放线菌素D法测定过表达SQLE在LOVO和SW1116细胞中抑制细胞凋亡(图3一).在表达SQLE的SW1116和LOVO细胞中,caspase-3、caspase-7和聚adp -核糖聚合酶(PARP)的裂解(活性)蛋白表达的减少进一步证实了SQLE对凋亡的抑制作用(图3 b).相反,SQLE敲低可促进HT29和DLD1细胞的凋亡(图3 c),同时增加裂解的caspase-3、caspase-7和PARP (图3 d).我们通过流式细胞术分析细胞周期分布,发现SQLE过表达LOVO和SW1116细胞增加S期细胞数量,同时G1期细胞数量减少(图3 e).与此相一致的是,SQLE过表达增加了增殖细胞核抗原(PCNA)、细胞周期蛋白D1和CDK4的蛋白表达,但抑制了细胞周期抑制剂p21的表达Cip1p27Kip1和p53 (图3 f).相反,在HT29和DLD1细胞中,SQLE敲除可诱导G1细胞周期阻滞(图3 g),表明SQLE加速了G1-S的进展。SQLE敲低抑制PCNA、cyclin D1和CDK4,促进细胞周期抑制剂的表达(图3 h).总的来说,SQLE通过抑制细胞凋亡和激活细胞周期进程来促进CRC的生长。

角鲨烯环氧化酶(SQLE)在结直肠癌(CRC)细胞中抑制细胞凋亡,促进细胞周期进展。(A)通过annexin V/7-氨基放线菌素D (7-AAD)染色和流式细胞术检测,SQLE过表达抑制LOVO和SW1116细胞凋亡。(B) Western blot分析表明,SQLE过表达抑制了裂解PARP、caspase-3和caspase-7的表达。(C)通过annexin V/7-AAD法抑制HT29和DLD1细胞SQLE诱导细胞凋亡。(D) SQLE敲除增加裂解PARP、caspase-3和caspase-7的表达。(E) SQLE在LOVO和SW1116细胞中过表达导致G0/ G1通过碘化丙啶(PI)染色和流式细胞术评估,S期细胞数量增加。(F) SQLE过表达增强了促进G1期进展的PCNA、cyclin D1和CDK4的表达,而抑制了G1守门人p21的表达Cip1p27Kip1和p53。(G) HT29和DLD1细胞中SQLE的敲除导致G细胞的积累0/ G1S期细胞数量减少。(H) SQLE沉默抑制PCNA、cyclin D1和CDK4,诱导p21Cip1p27Kip1和p53表达。两组间的差异由双尾Student 's t检验决定。* P < 0.05;* * p < 0.01。所有实验均进行3次,每组3个重复。
结肠特异性转基因SQLE在小鼠中的表达加速结肠肿瘤的发生
为了确定SQLE在结直肠肿瘤发生中的相关性,我们通过将Rosa26-Sqle tg小鼠与Cdx2-Cre杂交构建结肠特异性SQLE tg小鼠ERT2老鼠(图4一).注射他莫昔芬可激活结肠特异性过表达Sqle,它可激活结肠上皮中的Cre the (图4 b).为了启动结直肠肿瘤的形成,我们每周向野生型和Sqle tg小鼠注射10 mg/kg AOM,共6周(图4 b).第一次注射AOM后180天,处死小鼠,分析结肠。所示图4 c与野生型小鼠相比,Sqle tg小鼠的肿瘤发生率(10/10 vs 6/10)、肿瘤数量(p<0.01)和肿瘤负担(p<0.01)均增加,提示Sqle在CRC肿瘤发生中具有致瘤作用。H&E染色组织学检查证实Sqle tg小鼠结肠形成CRC,结肠发育不良和重度发育不良小鼠比例增加(图4 d).与这些发现一致的是,SQLE在结直肠癌中的致癌功能可由Ki-67评分所证实的细胞增殖增加(图4 e)但通过末端脱氧核苷基转移酶dUTP缺口末端标记(TUNEL)评分降低细胞凋亡(图4 f)与野生型小鼠比较。为了识别Sqle tg小鼠中差异表达的基因,我们对Sqle tg小鼠及其野生型窝鼠的结肠组织进行了癌症路径PCR分析。细胞周期进展标志物包括Mki67、Ccnd2和E2f4为最高上调基因,而促凋亡基因Casp9和Casp2下调,证实了Sqle在CRC肿瘤发生中促进细胞增殖和抑制凋亡的作用(图4 g).

大肠特异性角鲨烯环氧化酶(Sqle)转基因(tg)表达促进小鼠结直肠肿瘤的发生。(A)结肠特异性条件Sqle tg小鼠的构建。将Sqle转基因引入到Rosa26基因座中,Loxp位点位于5 '端STOP密码子的两侧。然后将这些小鼠与Cdx2-Cre交叉ERT2获得结肠特异性Sqle tg小鼠。6周龄时给予他莫昔芬注射液(100 mg/kg,腹腔注射)4次(1次/天)。Western blot分析证实SQLE在结肠组织中过表达。(B) SQLE促进化学诱导结直肠癌(CRC)。6周龄Sqle tg小鼠及野生型(WT)胎鼠(每组10只)腹腔注射他莫西芬(100 mg/kg,腹腔注射4次),随后再注射氮氧甲烷(AOM)注射液(10 mg/kg,腹腔注射)6次(每周1次)。第24周时处死小鼠。(C)结肠特异性Sqle过表达显著增加结肠肿瘤数量和负担(每组n=10)。(D)由病理学家进行的组织学检查证实了SQLE促进了肿瘤的发展,因为在SQLE tg小鼠中高度不典型增生的比例为70%,而在WT小鼠中相应的比例为20%。(E) Ki-67染色显示,Sqle tg小鼠结肠组织细胞增殖能力高于WT小鼠。(F) TUNEL染色显示,与WT小鼠相比,Sqle tg小鼠结肠细胞凋亡减少。 (G) Cancer Pathway PCR array analysis of colon tissues from Sqle tg and WT mice, which were first injected with tamoxifen (100 mg/kg, intraperitoneally, 4 times) and then sacrificed after 3 months. (H) The construction of CRISPR/Cas9-induced whole body Sqle knockout (KO) mice. Downregulation of SQLE in heterozygous Sqle KO mice colon was confirmed by western blot analysis. (I) Sqle KO mice and WT littermates were subjected to AOM-DSS regimen to induce CRC. Sqle KO mice developed significantly less colon tumours as compared with WT mice (n=5 in each group). The difference between two groups was determined by two-tailed Student’s t-test.
SQLE敲除小鼠可抑制结直肠肿瘤的发生
接下来,在基于CRISPR/ cas9的全身SQLE KO小鼠中测定了SQLE对CRC发育的影响。我们产生杂合子Sqle小鼠(图4 h),并进行AOM-DSS化学致癌模型(图4我).所示图4我与野生型窝鼠相比,Sqle KO小鼠结肠肿瘤数量减少(p<0.05)。总之,我们在Sqle tg小鼠和Sqle KO小鼠中的研究结果都证实了Sqle在结直肠肿瘤发生过程中的致癌功能。
SQLE促进胆固醇从头合成
SQLE是胆固醇生物合成的二级限速酶。因此我们评估了它对胆固醇生物合成途径的影响。在LOVO和SW1116细胞中,SQLE过表达增加胆固醇生物合成基因(HMGCS1、HMGCR、MVK、PMVK、MVD、IDI1、FDPS和FDFT1) mRNA的表达(在线补充图S2),而SQLE敲除则产生相反的效果(在线补充图S3).Western blot分析显示,SQLE过表达细胞或SQLE tg小鼠HMGCR、FDFT1和FDPS蛋白水平升高,而SQLE低表达或SQLE小鼠+ /−老鼠降低了这些基因的蛋白质水平(在线补充图S2和S3).这些数据表明,SQLE促进胆固醇从头合成途径,这有助于增加CRC细胞的细胞增殖。
Sqle转基因小鼠出现肠道失调
除了SQLE对CRC细胞的内在作用外,我们还询问了SQLE是否对结肠微环境有影响。肠道菌群失调是结直肠肿瘤发生的一个新病因。19因此,我们想知道在体内,肠道失调(至少部分)是否介导了Sqle的致癌作用。对3月龄时从Sqle tg和野生型小鼠收集的粪便进行鸟枪宏基因组分析。beta多样性分析没有显示出Sqle tg小鼠与野生型小鼠的单独聚类(图5一个),而Sqle tg的alpha多样性指数呈下降趋势(p=0.093) (图5一个).我们在物种水平上测定了Sqle tg和野生型小鼠的细菌组成的变化,并揭示了两组间具有显著丰度差异的异常细菌种类(fold change >2;p < 0.05) (图5 b).有趣的是,我们观察到明显富集的致病菌种类,如脱磷孤菌属fairfieldensis,红球菌属erythropolis,流产布鲁氏菌而且衣原体muridarum在Sqle tg小鼠体内富集,而保护细菌则减少,包括链霉菌属violaceusniger而且假单胞菌spLeaf58(图5 c).这些关键菌种的变化经qPCR (图5 d).这些结果表明,Sqle在体内过表达可促进肠道失调。

结肠特异性角鲨烯环氧化酶(Sqle)转基因(tg)表达在小鼠肠道菌群和代谢产物失调。(A)在注射他莫昔芬3个月后,对Sqle tg小鼠和野生型(WT)同胎小鼠的粪便进行宏基因组测序。左图为Sqle tg (n=8)和WT (n=9)小鼠间细菌组成的主成分分析(PCA)。Sqle tg小鼠的α多样性呈下降趋势(右)。(B和C)火山图和热图分析发现29种富集菌,包括致病菌,脱磷孤菌属fairfieldensis,红球菌属erythropolis,流产布鲁氏菌而且衣原体muridarum,以及34个耗尽的细菌如链霉菌属violaceoruber,假单胞菌spLeaf58在Sqle tg小鼠中。(D) Sqle tg小鼠体内关键菌种改变的定量PCR (qPCR)验证。(E)对3月龄Sqle tg小鼠和WT同窝小鼠的粪便样本进行全球代谢组学分析。代谢物谱的PCA分析(每组n=9)。(F和G)火山图和热图分析发现9种富集的代谢物,如脱氧胆酸(DCA)、岩石胆酸(LCA)、牛磺脱氧胆酸(TDCA)和鹅脱氧胆酸(CDCA);加上64个消耗的代谢产物Sqle tg小鼠与WT的窝鼠相比。(H)二级胆汁酸的液相色谱-质谱靶向代谢组学分析。* P < 0.05;* * p < 0.01。
Sqle转基因小鼠出现肠道代谢组学改变
由于已知一些富集的致病菌种类具有不同的代谢活性,我们接下来对相同的粪便样本进行了非靶向代谢组学分析。主成分分析表明,Sqle tg的代谢组可与野生型小鼠的代谢组分离(图5 e).专注于差异丰富的代谢物(图5 f, G),我们发现次生胆汁酸,如石胆酸、鹅脱氧胆酸、牛脱氧胆酸和脱氧胆酸在Sqle tg小鼠的粪便中较野生型小鼠富集(图5克).通过靶向代谢组学分析(图5 h).
由于次生胆汁酸是通过肠道微生物群的作用从初级胆汁酸转化而来的,20.我们研究了Sqle tg小鼠肠道细菌种类和代谢物之间的相关性,显示差异丰度(图6).我们观察到次生胆汁酸与c . muridarum这意味着它可能有助于增加次生胆汁酸。此外,所有四种致病菌均与关键的代谢物改变呈正相关(图6).因此,Sqle转基因表达调节小鼠结直肠癌发展中的肠道微生物群和代谢物。

在转基因角鲨烯环氧氧化酶(Sqle)小鼠和接受Sqle tg小鼠粪便菌群移植(FMT)的无菌小鼠中,肠道微生物-代谢物轴有助于肠道屏障功能障碍。(A)相关分析显示致病菌与次生胆汁酸等代谢产物的改变呈正相关。(B)注射他莫昔芬3个月后,Sqle tg小鼠和野生型(WT)胎鼠进行异硫氰酸荧光素(FITC)-葡聚糖肠通透性试验,结果显示Sqle tg小鼠肠道屏障受损。(C)电镜证实Sqle tg小鼠肠道紧密连接受损。(D) Western blot分析显示紧密连接蛋白Jam-c和occludin在Sqle tg小鼠中表达下调。(E) Sqle tg小鼠Muc2信使RNA (mRNA)表达下调。(F)从Sqle tg和WT窝鼠中收集的粪便灌胃给6周龄的无菌小鼠。1个月后,小鼠献祭前4小时灌胃fitc -葡聚糖进行通透性测定。无菌小鼠灌胃Sqle tg小鼠粪便后,肠道通透性明显增强。(G)电子显微镜证实用Sqle tg小鼠粪便喂养的肠道紧密连接受损无菌小鼠。 (H) Western blot analysis showed that tight junction proteins Jam-c and occludin were downregulated in germ-free mice gavaged with stool from Sqle tg mice. (I) Germ-free mice transplanted with Sqle tg mice stool showed increased Ki-67 staining in the colon epithelium as compared with those gavaged with WT mice stool. Two-tailed Student’s t-test was used for the difference determination between two groups.
SQLE转基因过表达会损害肠道屏障功能,促进炎症反应
次生胆汁酸是已知的脂毒素,其水平升高是CRC的重要危险因素。21它们对结肠上皮细胞有毒,已知会破坏肠道屏障功能。22肠道微生物失调也与肠道屏障的破坏密切相关。23因此,我们用fitc -葡聚糖经上皮通透性试验比较了Sqle tg小鼠和野生型凋落鼠的屏障功能。灌胃fitc -葡聚糖后,Sqle tg小鼠血清中fitc -葡聚糖含量高于野生型小鼠(图6 b).电子显微镜(EM)证实Sqle tg小鼠的紧密连接被破坏(图6 c).上皮紧密连接由细胞间连接蛋白复合物维持。我们通过western blot检测关键紧密连接蛋白的表达,发现与野生型小鼠相比,Sqle tg小鼠结肠中Jam-c和occludin表达下调(图6 d).此外,Sqle tg小鼠内粘液的关键成分Muc2在mRNA水平上被下调(图6 e).“渗漏”肠道的一个主要后果是诱导促炎症反应。与此观点一致的是,对Sqle tg结肠组织的炎症途径PCR阵列分析显示,促炎因子Cxcl3、Crp和Cxcr2 (在线补充图S4).这些结果表明,在Sqle tg小鼠中,肠道代谢异常-代谢物轴可能与肠道屏障功能障碍有关。
SQLE过表达诱导的肠道菌群失调促进无菌小鼠肠道屏障功能障碍和结肠上皮细胞增殖
为了研究Sqle tg表达诱导的肠道失调是否对结肠细胞增殖有直接影响,从AOM模型中收集的Sqle tg小鼠和野生型小鼠的粪便被移植到无菌(GF)小鼠。一个月后,小鼠被处死。不同粪便灌胃对体重无影响。在献祭前,我们进行了fitc -葡聚糖经上皮通透性试验,结果显示,用Sqle tg小鼠粪便灌胃的GF小鼠血清中fitc -葡聚糖含量显著高于用野生型小鼠粪便灌胃的GF小鼠(图6 f)及肠屏障破坏经电子显微镜证实(图6克).与野生型粪便相比,Sqle tg粪便灌胃的GF小鼠中连接粘附蛋白Jam-c和occludin均下调(图6 h).Ki-67染色结果显示,与野生型小鼠相比,Sqle tg小鼠粪便显著促进了GF小鼠结肠上皮细胞的增殖(图6我).这些数据强调了sqle诱导的肠道菌群失调在损害肠道屏障功能和促进结肠细胞生长方面的功能作用。
Sqle的药理抑制作用可提高结直肠癌化疗的疗效
鉴于SQLE在结直肠癌中具有致癌功能,我们下一步研究SQLE能否在结直肠癌细胞中靶向治疗。为了测试SQLE是否代表CRC的可行治疗靶点,我们用fda批准的真菌SQLE抑制剂特比萘芬治疗HT29和DLD1细胞。所示图7在体外,特比萘芬剂量依赖性地抑制HT29和DLD1细胞的生长。除结直肠癌细胞系外,特比萘芬显著损害原代结直肠癌类器官的生长(图7).接下来,我们评估了特比萘芬与奥沙利铂或5-FU(用于治疗结直肠癌的常见化疗药物)联合使用的效果。我们在存在或不存在特比萘芬的情况下进行了细胞活力测定,结果显示特比萘芬使HT29和DLD1细胞对奥沙利铂和5-FU的生长抑制作用敏感(图7 b).联合指数(CI)定量评估显示特比萘芬联合奥沙利铂(CI: 0.39-0.63)或5-FU (CI: 0.332-0.472)具有协同作用(<1)(图7 c).细胞凋亡实验结果表明,特比萘芬联合奥沙利铂和5-FU在体外协同诱导细胞凋亡(图7 d).特比萘芬也增强了5- fu诱导的细胞周期阻滞(在线补充图S5).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
角鲨烯环氧化酶(SQLE)是结直肠癌(CRC)的治疗靶点。(A)特比萘芬,一种食品和药物管理局批准的靶向SQLE的药物,剂量依赖性地抑制HT29和DLD1细胞的细胞活力,以及原发性CRC类器官。(B)特比萘芬联合治疗使HT29和DLD1细胞对5-氟尿嘧啶(5-FU)或奥沙利铂的生长抑制作用敏感。(C)联合指数(CI)显示特比萘芬+5-FU或奥沙利铂对CRC细胞具有协同作用,CI <1。(D)细胞凋亡实验显示特比萘芬+5-FU或奥沙利铂(48小时)协同诱导CRC细胞凋亡。(E)裸鼠皮下注射HT29细胞,随机分组,然后用(1)对照剂,(2)特比萘芬(80 mg/kg,每日口服),(3)5- fu (30 mg/mg,腹腔注射,每周一次),(4)奥沙利铂(10 mg/kg,腹腔注射,每周两次),(5)特比萘芬+5- fu和(6)特比萘芬+奥沙利铂治疗。特比萘芬单独治疗可显著抑制肿瘤生长~50%。另一方面,与5-FU或奥沙利铂单独治疗相比,特比萘芬联合5-FU或奥沙利铂在抑制裸鼠肿瘤大小方面表现出协同效应。通过重复测量方差分析确定肿瘤生长速率差异的显著性。* * p < 0.01; ***p<0.001; ****p<0.0001. All in vitro experiments were conducted 3 times in triplicate.
为了证实特比萘芬联合5-FU或奥沙利铂抑制结直肠癌生长的协同作用,我们在裸鼠皮下建立了异种移植模型。HT29细胞(1×106)植入裸鼠背侧,分为6组:(1)载药组;(2) terbinafine;(3)研究者用;(4)铂;(5)特比萘芬+5- fu和(6)特比萘芬+奥沙利铂。所示图7 e单药治疗抑制肿瘤生长约50% (p<0.01)。重要的是,与对照剂相比,特比萘芬+5-FU或特比萘芬+奥沙利铂的联合治疗均可引发完全生长停滞(p<0.001),且明显优于单一药物治疗(p<0.01)。我们的数据表明,SQLE在结直肠癌中是一个可操作的靶点,抑制SQLE可以提高结直肠癌化疗的疗效。
讨论
在本研究中,我们利用细胞系、结肠特异性SQLE tg小鼠和SQLE KO小鼠模型建立了SQLE作为结直肠癌的致癌因子。我们发现,SQLE通过细胞内在效应和肠道微生物-代谢产物轴的调节调节肿瘤的发生,分别驱动肿瘤细胞增殖和肠屏障功能障碍。我们进一步证明,SQLE抑制剂特比萘芬可通过与化疗联合发挥协同效应,重新用于CRC的治疗。与我们的临床前研究结果相印证的是,SQLE在CRC中过表达,其表达预示患者生存期较差,暗示SQLE是CRC的治疗靶点和预后因素。
我们的工作清楚地表明,SQLE在CRC中作为肿瘤细胞固有的致癌因子。在CRC细胞中异位表达SQLE可通过加速细胞周期进程和抑制细胞凋亡促进细胞增殖,而SQLE的KO则具有相反的作用。为了验证SQLE在体内的作用,我们建立了结肠特异性SQLE tg小鼠和SQLE+ /−综合评价Sqle功能获得和功能丧失在结直肠癌发生中的作用。与我们认为SQLE是一种致癌基因的观点一致,结肠特异性SQLE过表达加重了aom诱导的CRC,无论是在肿瘤发生率和疾病严重程度方面,SQLE tg小鼠的肿瘤均表现出显著较高的细胞增殖指数。相反,Sqle KO损害了aom - dss诱导的CRC。SQLE的促肿瘤作用可能部分归因于诱导胆固醇生物合成,这是支持细胞增殖的关键。所有这些发现共同表明SQLE在CRC肿瘤发生过程中起致癌基因的作用。
越来越多的证据表明,结直肠肿瘤的发生涉及肿瘤细胞及其微环境的复杂相互作用。24特别是,肠道菌群越来越被认为是结直肠癌的诱因。在此之前,我们已经证明用结直肠癌患者的粪便灌胃GF小鼠可以显著诱导结肠细胞增殖和肿瘤发生。19然而,目前尚不清楚基因改变和肠道失调之间的相互作用是否在结直肠肿瘤的发生中起作用。在这里,我们证明Sqle转基因在小鼠体内的表达可诱导肠道失调,其特征是几种致病菌的富集,包括d . fairfieldensis,r . erythropolis,b .流产而且c . muridarum,它们与各种病理有关25日- 27日;虽然具有抗炎和抗肿瘤特性的潜在保护细菌,美国violaceusniger而且假单胞菌spLeaf58耗尽。28 - 30与野生型小鼠相比,将Sqle tg粪便移植给GF小鼠显著促进结肠上皮细胞增殖,提示Sqle诱导的肠道失调在CRC中起偶然作用。这些数据表明,Sqle对肠道微生物群的影响超出了它对肿瘤细胞增殖和存活的直接影响。
研究表明,肠道内代谢物和菌群可以相互影响。31代谢组学分析显示Sqle tg小鼠粪便中次生胆汁酸的诱导。由于次生胆汁酸是微生物代谢的产物,我们综合分析了宏基因组和代谢组数据集。事实上,我们发现次生胆汁酸与c . muridarum是Sqle tg小鼠的一种异常致病菌。次生胆汁酸是一种重要的脂毒素,已知可诱导结肠上皮细胞DNA损伤32促进小鼠结肠癌的形成。33与此一致的是,我们发现Sqle tg小鼠的肠道屏障功能受损,表现为肠道通透性增加,EM分析和紧密连接蛋白下调。在Sqle tg小鼠中,伴随肠道屏障完整性受损的是炎症反应,这进一步加剧了结直肠肿瘤的发生。这些结果表明,肠道失调-代谢-肠道屏障轴可能在一定程度上促进了SQLE在体内的肿瘤促进作用。
鉴于SQLE在CRC发展中的重要作用,我们接下来评估了它作为治疗靶点的潜力。因此,我们测试了特比萘芬,一种fda批准的抗真菌感染药物,作为一种新的CRC治疗方法。与SQLE的致癌作用一致,特比萘芬对SQLE的阻断作用对CRC细胞产生生长抑制作用。特别是特比萘芬与结直肠癌常用化疗药物5-FU或奥沙利铂联合,可协同抑制结直肠癌的体外和体内生长。我们的研究结果表明,特比萘芬的重新使用可能是联合化疗治疗结直肠癌的一种策略。
与我们的实验结果相一致的是,对独立的人类结直肠癌队列的分析显示,SQLE在结直肠癌中过表达,其表达预示着结直肠癌患者的生存期较差。与我们的数据一致,金等和他等两者都报道了SQLE蛋白表达预示结直肠癌患者生存期较差。10 11恰恰相反,君等研究表明,高SQLE预示着良好的预后,特别是在IV期结直肠癌患者中,SQLE的缺失促进了结直肠癌细胞的转移表型。因此,SQLE可能对结直肠癌的生长和转移有不同的影响。未来利用SQLE转基因或KO小鼠研究SQLE在结直肠癌转移中的作用是有必要的。
总之,我们的工作确立了SQLE在CRC中的致癌作用。SQLE促进CRC细胞增殖和存活,并通过引起肠道菌群和肠道屏障功能障碍对肿瘤微环境产生影响。特比萘芬靶向SQLE显著提高了常规化疗的疗效,提示其在未来的CRC治疗中具有潜在的临床应用价值。
数据可用性声明
根据合理的要求提供数据。所有资料均可向通讯作者索取。
伦理语句
病人同意发表
伦理批准
本研究涉及人类参与者,并获得了北京大学肿瘤医院人类伦理委员会和香港中文大学(ID: CREC Ref. 2019.425)的批准。参与者在参与研究前给予知情同意。所有动物研究均按照香港中文大学动物实验伦理委员会(amuweco -20192185)批准的指南进行。
参考文献
补充材料
脚注
CL和YW是共同第一作者。
调整通知这篇文章在Online First发布后进行了修改。在线补充文件1已被替换。表S4中的Human SQLE qPCR引物校正如下:Human SQLE qPCR Forward: 5 ' -GATGATGCAGCTATTTTCGAGGC-3 ';反向:5 ' -CCTGAGCAAGGATATTCACGACA-3。
贡献者李传根(CL)进行了实验,分析了数据并起草了手稿;YW为动物工作提供设施;DL和YZ提供了转基因小鼠模型并对研究进行了评价;CCW对研究和修改稿进行了评论;OOC和Changan Liu (CL)进行了生物信息学分析;YL进行了类器官实验;XZ和JJYS对研究进行了评论;WK和KFT辅助组织芯片;JY设计,监督项目,修改手稿。JY为本论文的担保人。
资金国家重点研发计划项目(No. 2020YFA0509200/2020YFA0509203)、国家自然科学基金(NSFC)资助;81972576)、香港研资会- crf (C4039-19GF及C7065-18GF)、香港研资会- grf(14110819、24100520及14108718)、健康及医学研究基金(06170686及07181256)及中大校长酌情基金。
相互竞争的利益没有宣布。
来源和同行评审不是委托;外部同行评议。
补充材料本内容由作者提供。它没有经过BMJ出版集团有限公司(BMJ)的审查,也可能没有经过同行评审。讨论的任何意见或建议仅仅是那些作者(s)和不被BMJ认可。BMJ放弃从放在内容上的任何依赖产生的所有责任和责任。如果内容包含任何翻译材料,BMJ不保证翻译的准确性和可靠性(包括但不限于当地法规、临床指南、术语、药品名称和药物剂量),并且不对翻译和改编或其他原因引起的任何错误和/或遗漏负责。
请求的权限
如果您希望重用这篇文章的任何部分或全部,请使用下面的链接,它将带您访问版权清除中心的RightsLink服务。您将能够快速获得价格和以多种不同方式重用内容的即时许可。