条文本

摘要
客观的凋亡脱落的增加与肠屏障功能障碍和炎症性肠病(IBD)的发展有关。相反,生理性细胞脱落允许上皮单分子层的更新而不损害屏障功能。在这里,我们研究了活细胞挤压在IBD上皮屏障改变中的作用。
设计利用肠上皮细胞中的条件GGTase和RAC1敲除小鼠(Pggt1b我ΔIEC而且Rac1我ΔIEC小鼠),活体显微镜,免疫染色,机械生物学,类器官技术和RNA测序,我们分析了肠上皮细胞脱落的改变。此外,我们检查了IBD患者的人体肠道组织和肠道类器官的细胞脱落改变和RAC1功能。
结果上皮Pggt1b缺失导致细胞骨架重排和紧密连接重分布,由于细胞脱落受阻而导致细胞过度拥挤,最终导致小肠上皮渗漏和自发粘膜炎症,在较小程度上发生在大肠。体内和体外研究(敲除小鼠,类器官)都确定RAC1是GGTase的一个靶点,主要参与烯丙化依赖的细胞骨架动力学、细胞力学和上皮细胞脱落。此外,IBD患者肠道组织的炎症区域表现出漏斗状结构,细胞脱落受阻的迹象和RAC1功能受损。RAC1在人肠道类器官中的抑制引起了肌动蛋白的改变,与阻止细胞脱落相容。
结论上皮RAC1功能受损导致细胞过度拥挤和上皮渗漏,从而诱发慢性肠道炎症。上皮RAC1是细胞骨架动力学、细胞力学和肠细胞脱落的关键调节因子。RAC1的调控可能用于IBD患者肠道上皮完整性的恢复。
- 肌动蛋白细胞骨架
- 炎症性肠病
- 上皮屏障
- 肠上皮细胞
- 紧密连接
数据可用性声明
所有与研究相关的数据都包含在文章中或作为补充信息上传。
这是一篇开放获取的文章,根据创作共用署名非商业(CC BY-NC 4.0)许可证发布,该许可证允许其他人以非商业方式分发、混音、改编、在此基础上进行构建,并以不同的条款许可其衍生作品,前提是正确引用原始作品,给予适当的荣誉,任何更改都已注明,并且使用是非商业性的。看到的:http://creativecommons.org/licenses/by-nc/4.0/。
数据来自Altmetric.com
本研究的意义
关于这个问题我们已经知道了什么?
上皮屏障功能有助于维持肠道组织稳态,避免肠道炎症。
病理性细胞脱落与慢性肠道炎症环境下上皮完整性的改变有关。
尽管生理性细胞脱落控制上皮细胞数量以维持上皮细胞的完整性,但其与炎症性肠病(IBD)发病机制的关系尚不清楚。
新的发现是什么?
GGTase和/或rac1缺陷上皮细胞的上皮内固有改变导致屏障破坏和肠道炎症。
在GGTase和缺乏rac1的上皮细胞中,细胞骨架重排和细胞力学的改变导致细胞脱落和细胞过度拥挤。
炎性炎症患者肠组织中生理细胞脱落和过度拥挤的停止迹象。
IBD患者肠组织上皮表面的上皮RAC1功能被特异性调节。
本研究的意义
在可预见的未来,它会对临床实践产生怎样的影响?
肠上皮细胞(IECs)中的RAC1功能可能被用于诊断和/或治疗IBD患者的上皮恢复。
上皮细胞力学改变可用于识别IBD中的上皮渗漏,甚至用于预测耀斑。
在IBD患者中,药物抑制RAC1可能对上皮完整性产生有害影响。
简介
在炎症性肠病(IBD)中,慢性肠道炎症是由于对微生物群的反应加剧。1 2因此,上皮渗漏是IBD患者的一个显著特征。3 4缓解期患者肠通透性增加5和亲戚,6 7以及影响上皮蛋白的单基因疾病中的ibd样表型,8 9在IBD发病过程中诱发上皮细胞破坏。
肠上皮细胞的更新需要在细胞挤压和增殖之间进行严格的调节,控制细胞数量和屏障的完整性。10细胞脱落与细胞死亡密切相关;Caspase激活发生在凋亡细胞挤压脱落之前,11或者由于活细胞脱离而失去生存信号。12同时,细胞脱落作为一种防御机制,以摆脱感染13或突变14细胞。根尖挤压缺陷导致了恶性肿瘤的特征,15基底挤压导致肿瘤扩散和转移。16
共聚焦显微镜研究表明,过度凋亡细胞脱落、通透性缺陷与肠道炎症之间存在相关性。4 17 18然而,生理性细胞脱落在IBD中的作用尚不清楚。关于压电驱动的力学感觉,月19 - 21日s1pr2介导的rho依赖肌动蛋白收缩性导致单个活细胞挤压。22日23日紧密连接(tight junction, TJs)的重新调节抑制了生理上的暂时性上皮渗漏24和病理11细胞脱落。因此,TJ组装和细胞骨架动力学控制肠道通透性,25它们的改变可能有利于屏障破坏和炎症。因此,在克罗恩病中观察到TJ链的连续性/数量的变化26还有溃疡性结肠炎。27此外,与细胞骨架功能相关的蛋白质上皮缺乏的小鼠引起了结肠炎的易感性增加。28 29
小Rho GTPases与细胞骨架密切相关,调节上皮屏障功能30.并在上皮间隙关闭中起重要作用。22 33节翻译后戊烯酰化允许小的gtpase锚定在细胞膜和它们的功能。我们之前证明了Rho蛋白ggtase1 -戊烯化、上皮完整性和炎症之间的联系。34在本研究中,我们发现GGTase1在IECs内的缺失表达,主要导致细胞骨架重排和apical junction complex (AJC)蛋白重分布,导致细胞脱落阻滞,导致上皮渗漏和局部免疫反应的激活。RAC1是ggtase的一个靶点,在上皮细胞骨架动力学和细胞力学中起重要作用,其在IBD中的改变在阻止细胞脱落和屏障功能障碍中起关键作用。这些发现为IBD中上皮细胞恢复的治疗调节开辟了新的途径。
结果
肠屏障改变与肠道病理动力学Pggt1b我ΔIEC老鼠
为了确定IECs内诱导抑制戊烯酰化的功能效应和分子事件序列,我们分析了IECs的肠道病理Pggt1b我ΔIEC随着时间的推移。删除Pggt1b基因在他莫西芬治疗2天后发生,导致GGTase1表达下降,非戊烯酰化蛋白在回肠组织和分离的IECs中积累(在线补充图1)。虽然我们在第6天(高分辨率内窥镜检查)没有检测到任何肠道组织改变的宏观迹象,但我们可以观察到在第4天和第6天之间发生的肠道通透性增加的趋势(图1一个)。因此,在第4天就可以通过活体显微镜检测到小肠内右旋糖酐的通过(图1 b)。相比之下,回肠和十二指肠的粘膜结构完全破坏只能在稍后的时间点看到,导致屏障功能明显抑制(第8天),而大肠只能观察到轻微的改变(图1 c;在线补充图1)。关于肠道内潜在的炎症反应Pggt1b我ΔIEC我们仅在晚些时候(第8天)观察到回肠和十二指肠中IL-6和TNF-α的表达增加,但在结肠中没有(图1 d;在线补充图1),同时渗透MPO+回肠第6天中性粒细胞已明显(图1 e;在线补充图1)。这些数据显示上皮细胞的固有现象Pggt1b缺失导致渗透性缺陷和中性粒细胞早期浸润,并支持了上皮屏障缺陷导致肠病理的最初假设,这是由于IECs中ggtase介导的戊烯酰化被废除。基于严重程度,我们在后续研究中重点研究了影响小肠的表型Pggt1b我ΔIEC老鼠。

随着时间的推移,IECs内的戊烯酰化受到抑制而导致的肠道疾病。(A)体内/体外组织完整性和体内肠通透性的评估。结肠和小肠组织内窥镜照片(左);经粘膜给药fitc -右旋糖酐(4 KDa);血清浓度(µg/mL)(右)。(B)使用腔内吖啶黄(绿色)和罗丹明B-右旋糖酐(红色)对小肠屏障功能进行活体显微镜分析的代表图片。(控制,n = 8;第2天,n=6;第4天,n=5;第6天,n=5; day 8, n=2). (C) Histology analysis of ileum tissue using H&E staining. Representative pictures (left), and corresponding score (right). (D) Gene expression of IL-6 and TNF-α in ileum tissue (RT-qPCR; six independent experiments). (E) MPO immunofluorescence staining in cross-sections from ileum. Representative pictures (left), and corresponding quantification (right). Data are expressed as box-plots (Min to Max); seven independent experiments, except where indicated. One-way ANOVA, Dunnett’s multiple comparisons test. *P≤0.050; ** P≤0.001. ANOVA, analysis of variance.
阻止细胞脱落和细胞过度拥挤Pggt1b我ΔIEC老鼠
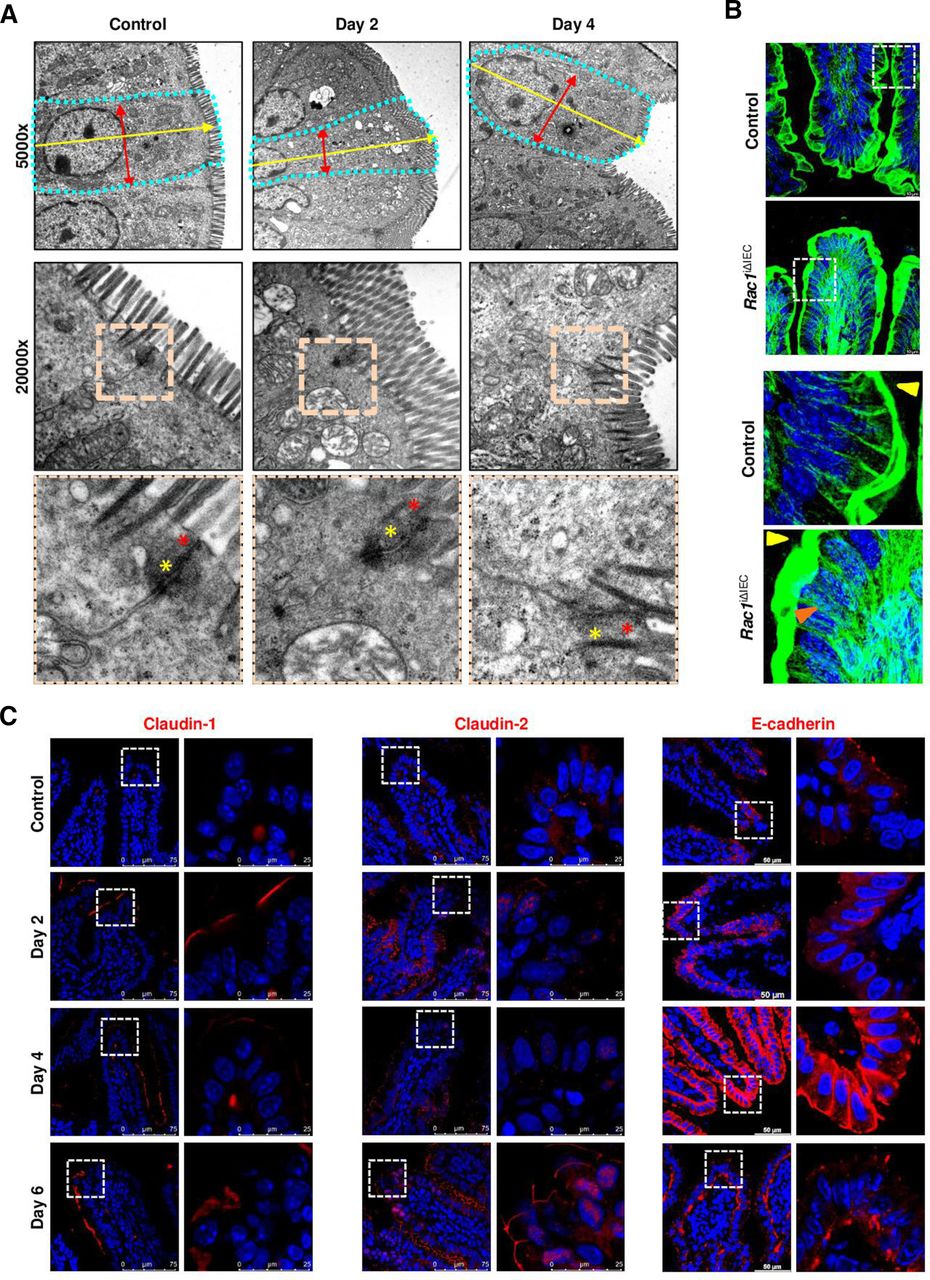
考虑到先前描述的肠道表型观察Pggt1b我ΔIEC老鼠,34我们分析了他莫西芬诱导的细胞脱落pggt1b-随着时间的推移而缺失,以识别早期上皮内固有的改变。利用活体显微镜,35我们可以证明,在治疗后的早期时间点,小肠细胞脱落率显著降低Pggt1b删除(第2天)(图2一个),与“渗透性iec”的初始累积相吻合(图2 b)。这些数据支持早期细胞脱落改变对小鼠肠道表型有重要影响Pggt1b我ΔIEC小鼠和指向受损或“阻止”细胞脱落。先前的出版物将漏斗状细胞的出现描述为由于邻近细胞基底侧压力而发生细胞挤压的早期事件,24这些漏斗状结构的积累可以解释为受损细胞挤压的替代品。除了E-cadherin和/或ZO-1的再分配,24通过肌动蛋白重分布(phalloidin染色)可识别漏斗状结构(在线补充图2)。有趣的是,在第2天回肠组织中出现漏斗状结构,显示部分膨出的IECsPggt1b删除(图2 c),与细胞停止脱落和完全脱落事件之间的转换有关(第2天和第4天)(在线补充图2)。部分挤压细胞的积累可能导致细胞过度拥挤,由于邻近细胞的压力增加而导致挤压和改变细胞形状。36因此,我们想知道我们是否可以观察到过度拥挤的上皮Pggt1b我ΔIEC老鼠。回肠IECs中ggtase介导的戊烯化抑制可观察到细胞密度(细胞/基底膜长度)增加(图2 c)。与此同时,细胞长度(第2天)和长度/直径比(第4天)显著增加,这与过度拥挤导致的细胞挤压一致(在线补充图2)。为了确认缺乏ggtase的IECs的形状改变,我们对回肠组织进行了透射电镜(TEM)分析。在他莫西芬缺失2天后,可以观察到细胞拉长(长度增加)和挤压(直径减小)(图3)。这与顶端肌动蛋白网络增大和AJC改变有关(图3)。综上所述,这些数据表明细胞脱落的早期阻滞Pggt1b导致细胞过度拥挤。

ggtase缺陷小肠的细胞脱落改变和过度拥挤(时间过程研究)。(A,B)使用腔内吖啶黄(绿色)和罗丹明B-右旋糖酐(红色)对细胞脱落进行活体显微镜分析。(控制,n = 8;第2天,n=6;第4天,n=5;第六天,n=5)。多重未配对t检验(A)时间序列代表性图片;白色椭圆表示细胞正在脱落;蓝绿色椭圆表示细胞脱落已完成,细胞位于管腔内,脱离上皮层。在某些情况下,由于组织收缩或蠕动,在图像采集过程中,根据焦平面的变化来校正z -位置。 Quantification of cell shedding rate considering exclusively events which are completed during the duration of the image acquisition; (number of cell shedding events/time/length) (top), and in a single villus (events/minute) (bottom), at a determined focus plane near the lumen (five villi/video; two videos/mouse). (B) Representative pictures of single villus from intravital microscopy experiment (left); orange arrows indicate permeable cells. Quantification of permeable cells (dextran is detected inside the cell) (events/villus; 10 villi/picture; two pictures/mouse). (C) F-actin fibre staining using AlexaFluor488-phalloidin in ileum tissue (green). Seven independent experiments. Mixed effect analysis. Representative pictures (left); quantification of funnel-like structures, indicated by white arrows (top right); quantification of cell density (number of cells/µm of basement membrane length) (bottom right). Data are expressed as box-plots (Min to Max). *P≤0.050; ***P≤0.0001.

对照组和对照组回肠组织透射电镜分析Pggt1b我ΔIEC小鼠(第2天和第4天)。细胞直径和细胞长径比的定量(左);代表性图片(右)。细胞形状由绿点线表示;红色箭头表示细胞直径(在两个侧膜之间),黄色箭头表示细胞长度(在基膜和顶膜之间)。在AJC内,红色星号表示紧密连接区,黄色星号表示粘附连接区。一个样本/组。数据以盒状图(最小值到最大值)表示。* P≤0.050。AJC,顶端结复合体;透射电子显微镜。
细胞骨架重排和细胞力学改变Pggt1b我ΔIEC老鼠
然后,我们的目标是描述这种受损细胞脱落背后的分子机制。生理细胞脱落的完成依赖于s1p介导的细胞通讯。22由于S1P2R在回肠组织中表达无明显改变Pggt1b我ΔIEC老鼠(在线补充图3),我们假设该信号检查点在烯丙化抑制时没有受损。接下来,上皮间隙的闭合取决于肌动蛋白的收缩能力,包括肌动蛋白的钱袋和从邻近细胞中爬行的细胞。37根据透射电镜分析(图3),高分辨率staris共聚焦显微镜证实回肠绒毛尖端IECs内肌动蛋白骨架发生改变Pggt1b我ΔIEC小鼠:顶端肌动蛋白网络增大,纤维重新分布(图4一)。然后,我们分析了肌球蛋白的结构,可以观察到,在对照组小鼠位于绒毛尖端的回肠iec中,MyosinIIA信号通常更强(图4 b),即发生生理细胞脱落的地方。但是,及早删除Pggt1b(2-4天),该信号均匀分布于绒毛,直至隐窝基底(图4 b)。总之,我们在IECs内发现了早期肌动蛋白重分布对烯丙化的抑制。

基因缺失后小肠细胞骨架重排及细胞力学改变Pggt1b。(A)回肠组织中使用AlexaFluor488-phalloidin染色f -肌动蛋白纤维(绿色)。高分辨率共聚焦显微镜(Leica Stellaris)。最大投影的代表图片(系统优化的z-stack)。黄色箭头表示顶端肌动蛋白网络;橙色箭头表示肌动蛋白纤维的再分布。三个独立的实验。(B)回肠组织肌球蛋白IIA染色(红色)的代表性图片(五次独立实验)。共聚焦显微镜(徕卡SP8)。(C)小肠IECs RT-FDC分析(n=4/组)。 (D) Expression and redistribution of selected candidate AJC proteins in ileum tissue (claudin-1, claudin-2 and E-cadherin). Immunostaining (top, red signal) and western blot (bottom). Band densitometry quantification (right). Minimum five independent experiments. Data are expressed as box-plots (Min to Max). One-way ANOVA, Dunnett’s multiple comparisons test. *P≤0.050. AJC, apical junction complex; ANOVA, analysis of variance; RT-FDC, real time fluorescence deformability cytometry.
由于细胞骨架重排和“挤压”细胞形状表明缺乏ggtase的IECs也改变了细胞力学,我们通过实时荧光变形细胞术(RT-FDC)分析了从小肠中新分离的IECs。38我们很快就发现了Pggt1b缺失(第3天),ggtase缺失IECs (EpCAM+CD45-)的特点是杨氏模量降低,表明变形能力增加(图4 c;在线补充图3)。这些观察结果与细胞内在现象(细胞骨架改变)一致,导致单个细胞力学特性的改变(RT-FDC数据),可能导致携带ggtase缺陷上皮细胞的小鼠上皮渗漏。
TJs蛋白的再分配显著地促进了细胞挤压时上皮层的封闭。11通过透射电镜观察到ggtase缺陷上皮细胞AJC的改变(图3),为了描述影响细胞脱落完成的细胞间连接蛋白的改变,我们在回肠组织中筛选了多达25种蛋白的表达。claudin蛋白(-1 ~ -2和-8)、occludin和ZO-1编码基因的调控表达;以及e -钙粘蛋白和β-连环蛋白(在线补充图3)提示TJ和AJ在缺失时可能发生变化Pggt1b。细胞间连接蛋白的功能主要取决于它们的亚细胞定位和与细胞骨架的相互作用。39事实上,在早期时间点(第2-4天),我们可以在绒毛尖端检测到claudin-2、claudin-8、claudin-18、E-cadherin和β-catenin的重新分布(信号增强),以及回肠横切面上claudin-1和ZO-1的染色减少;claudin-2、claudin-8和ZO1可通过WB证实调控蛋白的表达(第3天)(图4 d;在线补充图3)。在AJC内发生的变化中Pggt1b缺失后,部分变化可恢复(4-6天),如ZO1下调或claudin-2上调,而其他变化随时间持续,如claudin-1减少或claudin-18、E-cadherin或β-catenin表达增加。事实上,E-cadherin的再分配证实了漏斗状结构的积累(在线补充图2)。总之,我们的数据显示了细胞骨架重排,细胞形状和力学改变以及AJC蛋白重分布Pggt1b我ΔIEC老鼠。
肠道病理Pggt1b我ΔIEC小鼠并非主要由caspase介导的病理性细胞脱落引起
上皮渗漏Pggt1b我ΔIEC小鼠可被认为是细胞骨架重排和TJ分解的结果。然而,可渗透iec的出现可能是由细胞死亡引起的。40 41测定细胞脱落是否阻滞Pggt1b我ΔIEC小鼠与细胞死亡调节有关,我们研究了其动力学Pggt1b删除。总细胞死亡(TUNEL)无显著变化+细胞)或裂解的caspase-3+回肠内观察到细胞Pggt1b我ΔIEC老鼠随时间的变化(图5一个)。尽管如此,当上皮完整性被破坏时,在后期时间点可以观察到死亡细胞积累的明显趋势。此外,死亡细胞内裂解caspase-3激活的分析表明,凋亡之间存在潜在的转移(TUNEL+Casp3+)和非凋亡死亡(TUNEL+Casp3-) (在线补充图4)。因此,我们可以观察到早期小肠IECs中caspase-3的切割减少Pggt1b删除(第3天)(图5 b, C)。这些观察结果表明,诱导细胞死亡(病理性细胞挤压)可能与导致肠道表型和细胞脱落改变的ggtase -戊烯酰化抑制的初始事件无关,但可能作为一种放大事件发生,在后期进一步加重组织损伤。然后,我们想知道坏死作为一种替代细胞死亡途径是否在IECs中缺乏GGTase的情况下起作用。然而,尽管诱导基因表达mlkl在回肠Pggt1b我ΔIEC老鼠随时间的变化(在线补充图4),我们无法检测到MLKL蛋白在早期时间点的表达增加(图5 b, C)。最近有研究表明,gasdermin - e依赖性焦性细胞死亡在细胞挤压中起相关作用。42考虑到与膜完整性破坏相关的其他替代细胞死亡途径,我们还分析了焦亡的激活41在小肠中Pggt1b我ΔIEC老鼠。Gasdermin-D和Gasdermin-E在GGTase缺失的任何时间点均未检测到受调控的基因表达(在线补充图4)。然而,晚期小肠组织破坏与IECs内Caspase-1和Gasdermin-D的裂解有关,这表明焦化细胞死亡的激活是次要机制(图5 d)。因此,在ggtase缺陷上皮细胞中,caspase-3表达受损或直接抑制细胞凋亡以及可能激活另一种细胞死亡途径(坏死和/或pyroptosis)可能被认为是细胞骨架重排的次要机制。考虑到细胞死亡和挤压之间的密切关系,细胞骨架重排与细胞力学改变以及细胞凋亡和/或焦亡调节之间的关系还需要进一步的研究。

细胞死亡活化对小肠上皮内戊烯酰化的抑制作用。(A) TUNEL(绿色)和cleaved caspase-3(红色)在对照和回肠横截面上的染色Pggt1b我ΔIEC不同时间点的老鼠。TUNEL的代表性图片(上)和量化(下)+(左)和Cl。Casp-3+/绒毛(右)。六个独立的实验。(B,C) Western blot分析cleaved caspase-3和MLKL。代表性印迹图(上)和频带密度定量图(下)。(B)回肠组织(Cl。Casp-3;n = 8,控制;n = 10,Pggt1b我ΔIEC) (MLKL;n = 6,控制;n = 7,Pggt1b我ΔIEC)。(C)小肠分离IECs (Cl。Casp-3;n = 8,控制;n = 10,Pggt1b我ΔIEC) (MLKL;n = 6,控制;n = 7,Pggt1b我ΔIEC)。(D)小肠IECs中Caspase-1和Gasdermin-D切割的检测(western blot分析)。代表性印迹图(左)和频带密度定量图(右)。四个独立的实验。数据以盒状图(最小值到最大值)表示。单向方差分析,Dunnett多重比较检验(A,D)。未配对t检验(B,C)。* P≤0.050。
除了细胞死亡和挤压外,肠道上皮细胞的更替还由隐窝中的干细胞增殖和体外培养细胞沿隐窝/绒毛轴的迁移决定。与细胞死亡激活和炎症反应相似,Ki67表达显著下调表明回肠细胞增殖下降,只有在组织破坏未完全时才能观察到(在线补充图4)。利用BrdU掺入试验,我们无法观察到小肠上皮细胞在回肠中与对照组和对照组有不同的迁移行为Pggt1b我ΔIEC老鼠(在线补充图4)。这些数据证实,在IECs内缺乏GGTase的小鼠中,细胞死亡、增殖和迁移的改变并不是导致肠道稳态破坏的主要机制。
烯丙化在细胞骨架/上皮细胞脱落调控中的作用
为了鉴定功能调控上皮GGTase失活的戊烯酰化靶点,我们通过质谱分析了膜结合蛋白(第6天)。在小肠IECs的膜部分,各种戊烯酰化gtpase下调Pggt1b我ΔIEC如RAP1A, RHOG, RAP1B和RAC1 (表1)。基于其作为细胞骨架调节剂的功能,以及在上皮伤口闭合过程中与RHOA的紧密联系,43RAC1是我们研究的一个有吸引力的候选对象。有趣的是,尽管蛋白表达没有改变,但RAC1在胞浆内的积累在缺失后不久就被注意到Pggt1b(第3天),提示该蛋白在小肠IECs中存在早期功能障碍(图6;在线补充图5)。相反,早期时间点RHOA未见变化。

RAC1在小肠IECs中的功能对于维持肠道稳态至关重要。(A) RAC1和RHOA在对照和小肠IECs中的亚细胞定位Pggt1b我ΔIEC他莫西芬治疗第3天的小鼠。细胞质蛋白通过离心梯度分离,小gtpase通过western blotting检测。代表性印迹图(左)和频带密度定量图(右)。3个实验(n=6,对照组;n = 8,Pggt1b我ΔIEC)。(中)。表型的Rac1我ΔIEC老鼠。(B-D)肠道病理和细胞脱落改变Rac1我ΔIEC老鼠。(B)回肠组织H&E染色组织学分析(第7天)。代表性图片(左)和组织学评分(右)(n=10,对照组;n = 6,Rac1我ΔIEC)。(C)体内肠通透性评价。经粘膜给药fitc -右旋糖酐;血清浓度(µg/mL) (n=6,对照;n = 11,Rac1我ΔIEC)。(D)在小肠中使用腔内吖黄碱(绿色)和罗丹明b -右旋糖酐(红色)进行细胞脱落的活体显微镜分析。代表图片(左);定量细胞脱落率(在确定的焦点平面上单个绒毛中随时间发生的细胞脱落事件的数量(事件/分钟/µm),以及可渗透细胞(细胞内检测到右旋糖酐)(事件/绒毛)。(n = 8,控制;N =3,第3天;n=6,第6天)。(E)时间课程研究。三个独立的实验。回肠组织中使用AlexaFluor488-phalloidin(绿色)染色f -肌动蛋白纤维。代表图片(左); and quantification of funnel-like structures, indicated by white arrows (% of total cell shedding events) (top right); quantification of cell length/diameter ratio (bottom right). One-way ANOVA, Dunnett’s multiple comparisons test or unpaired t test. *P≤0.050; **P≤0.001; ***P≤0.0001. ANOVA, analysis of variance.
为了确定RAC1作为GGTase靶点在上皮完整性中的作用,我们在IECs中生成了诱导性条件敲除RAC1的小鼠(Rac1我ΔIEC老鼠)(在线补充图5)。引人注目的是,这些新生成的小鼠表现出与Pggt1b我ΔIEC有肠道病变的小鼠,体重减轻,组织结构破坏,尤其影响小肠,结肠则温和得多(图6 b;在线补充图5)。Rac1我ΔIEC小鼠出现屏障功能破坏(图6 c, D),回肠组织中促炎细胞因子(即TNF-α)的表达失调(在线补充图5)、细胞脱落率早期下降及可渗透细胞在小肠内积聚(图6 d)。在机制上,细胞死亡激活、细胞增殖下降或细胞沿回肠隐绒毛轴迁移改变可能是抑制大肠杆菌的主要关键机制Rac1IECs内的表达式(在线补充图5)。值得注意的是,我们还可以观察到caspase-3在Rac1我ΔIEC小鼠(第3天),但回肠和小肠IECs内MLKL表达无调节(在线补充图5)。随着时间的推移,我们无法在RAC-1缺陷的iec中检测到焦性细胞死亡的迹象(在线补充图5)。相反,在回肠可以观察到细胞脱落、阻滞和过度拥挤的迹象Rac1我ΔIEC老鼠(图6 e;在线补充图5)。回肠IECs透射电镜分析证实AJC上皮细胞形状拉长及改变(图7)。我们也观察到肌动蛋白纤维的重新分布(图7 b)以及肌萎缩症从绒毛顶端向下至隐窝底部的细胞质积聚和再分布(在线补充图5),模拟细胞骨架重排Pggt1b我ΔIEC老鼠。根据AJC的改变,早期删除rac1, claudin-1、claudin-2、claudin-8、E-cadherin、ZO-1、β-catenin表达增加,而claudin-18在回肠IECs中由膜向细胞核转移(图7 c;在线补充图5)。总之,RAC1是IECs中GGTase的一个重要靶点,用于导致细胞脱落阻滞的早期细胞骨架改变。

rac1缺陷肠上皮细胞骨架重排(A)回肠组织的电镜分析。代表性的图片。细胞形状由绿点线表示;红色箭头表示细胞直径(在两个侧膜之间),黄色箭头表示细胞长度(在基膜和顶膜之间)。在AJC内,紧密连接区由红色星号表示;黄色星号表示粘附体交界区。一个样本/组。(B)用AlexaFluor488-phalloidin染色f -肌动蛋白纤维(绿色)。高分辨率共聚焦显微镜(Leica Stellaris)。 Maximum projection (system optimised z-stack). Yellow arrows indicate apical actin network; orange arrows indicate actin fibres. (C) Detection of selected candidate AJC proteins in ileum tissue (claudin-1, claudin-2, E-cadherin). Immunostaining (red signal). Three independent experiments. AJC, apical junction complex.
依赖rac1的上皮内在机制
为了描述rac1依赖的上皮内在机制,我们利用小肠类器官培养。缺乏ggtase和缺乏rac1的类器官(在线补充图6)显示细胞活力下降(图8),表明尽管缺乏上皮外源性介质,上皮细胞仍发生了破坏。我们想知道干扰细胞死亡是否会损害体外上皮细胞的破坏。在GGTase和rac1缺陷类器官中,caspase和necroptosome抑制剂(Z-VAD+Necrostatin-1)的联合治疗可以延迟上皮损伤,但不能完全挽救,而后者也单独对这些抑制剂有反应。使用焦亡抑制剂双硫仑治疗未观察到任何效果(在线补充图6)。根据我们在体内的数据(图5),这些结果证实了细胞死亡诱导(凋亡和/或坏死)参与了GGTase或RAC1抑制导致的上皮破坏,但不是主要机制。引人注目的是,抑制GGTase-prenylation或RAC1的细胞骨架重排也发生在类器官中。肌动蛋白纤维在侧膜和基膜上的重分布伴随着漏斗状结构的积累(图8 b)。此外,在类器官中缺乏GGTase或RAC1时,肌球蛋白IIA向细胞质转移(在线补充图6)。我们观察到戊烯化抑制后claudin-2、claudin-18和β-catenin表达增加,而E-cadherin表达降低(图8 c)。在rac1缺陷类器官中,我们还证实claudin-2和β-catenin表达增加,E-cadherin和ZO-1表达减少,而claudin-1和claudin-18没有改变(图8 d)。在尖向外的类器官中也观察到肌动蛋白纤维的重新分布,在ggtase缺乏的类器官中也观察到漏斗状结构的积累(图8 e;在线补充图6)。然后,我们想知道这些类器官的细胞力学是否也发生了变化。我们利用3D牵引力显微镜来量化胶原-1凝胶中有机生成的收缩力(收缩力或收缩压力),测量力诱导的基质变形。44 45代表性控制类器官向内的基质变形稳定增大,表明力增大;而代表Pggt1b我ΔIEC而且Rac1我ΔIEC类器官显示向外变形,表明力松弛(图9 a - c;在线补充图6)。这种分化行为开始于他莫西芬治疗后约48小时,此时靶蛋白缺失,细胞表型开始改变(在线补充图6)。总之,这些数据表明GGTase-和rac1缺陷上皮细胞(类器官培养)的细胞力学发生了改变。

小肠类器官上皮细胞的内在机制。由小肠隐窝产生的缺乏ggtase和缺乏rac1的类器官。(A) PI掺入测定的细胞活力(红色)。代表性图片(左),相应的量化(死亡类器官百分比)(右)。(Pggt1b,四次实验;Rac1, 3次实验)。(B)用AlexaFluor488-phalloidin染色f -肌动蛋白纤维(绿色)(Pggt1b,五项实验;Rac1,四个实验)。白色箭头表示漏斗状结构或停止细胞脱落事件。(C, D)。免疫染色分析AJC候选蛋白。z-stack的最大投影(系统优化)。(C)缺乏ggtase的类器官。四个实验;除β-catenin和Claudin-1外,共3次实验。(D)缺乏rac1的类器官。三个实验; except for claudin-2, five experiments. (E) Apical-out GGTase-deficient organoids, representative pictures of Phalloidin staining. One experiment. Data are expressed as box-plots (Min to Max). Paired t-test. *P≤0.050. AJC, apical junction complex.

三维牵引力显微镜观察对照组、GGTase和rac1缺陷小肠类器官。(A)延时成像47小时至72小时之间(胶原蛋白聚合完成后48-73小时)的类器官生成的基质变形。代表性的控制类器官表现为向内的矩阵变形,表示力的增加,而代表性的Pggt1b我ΔIEC而且Rac1我ΔIEC类器官显示向外变形,表明力松弛。(B)控制的平均收缩力随时间变化,Pggt1b我ΔIEC而且Rac1我ΔIEC类器官,每一对都来自同一个重复实验。灰线表示下图中报告部队发展情况的47小时和72小时时间点。测量Pggt1b我ΔIEC相应的控制类器官进行了超过90小时,以证明力的趋势继续。(C) 47小时至72小时之间收缩力的相对和绝对变化以及收缩压力的绝对变化。每个点代表来自单个类器官的数据,颜色代表四个生物重复。黑色圆圈代表每个生物复制的平均值,黑色箭头代表所有类器官的平均值。P值由假设方差不相等的双向学生t检验计算。配对t检验。* P≤0.050。
我们的类器官数据显示,在缺乏非上皮因子的系统中,GGTase的上皮内禀改变或RAC1缺失会导致细胞骨架重排和上皮完整性的改变。为了更深入地了解这些上皮改变,我们对缺乏rac1的小肠类器官进行了RNASeq分析,并将其与野生型类器官进行了比较(在线补充图6)。在缺乏rac1的类器官中,有137个基因存在差异表达。通过RT-qPCR分析确定候选基因的调控表达(Adm2,Hif3a,Cxcl10,Klf2),在缺乏rac1和缺乏ggtase的类器官中。基因本体注释提示,上皮RAC1缺失的转录组改变与对细菌、机械刺激反应和免疫反应的改变有关。在缺乏rac1的类器官中,细胞表面、细胞膜、肌动蛋白骨架和顶端细胞间连接发生改变(在线补充图6)。在通路水平上(KEGG注释),rac1缺陷上皮细胞中调控了几种炎症相关通路,如TNF、NFkB、MAPK、TLR和IL17信号通路、细胞粘附和白细胞迁移或紧密连接。此外,GSEA研究表明,细胞黏附、趋化因子活性和TNF/IL17信号在rac1缺陷上皮细胞中被调节。RNASeq分析证实了我们提出的机制:细胞骨架改变导致顶端肌动蛋白重排和AJC重分布,会导致上皮渗漏和对微生物群或机械刺激的免疫反应改变。
IBD中的细胞脱落改变
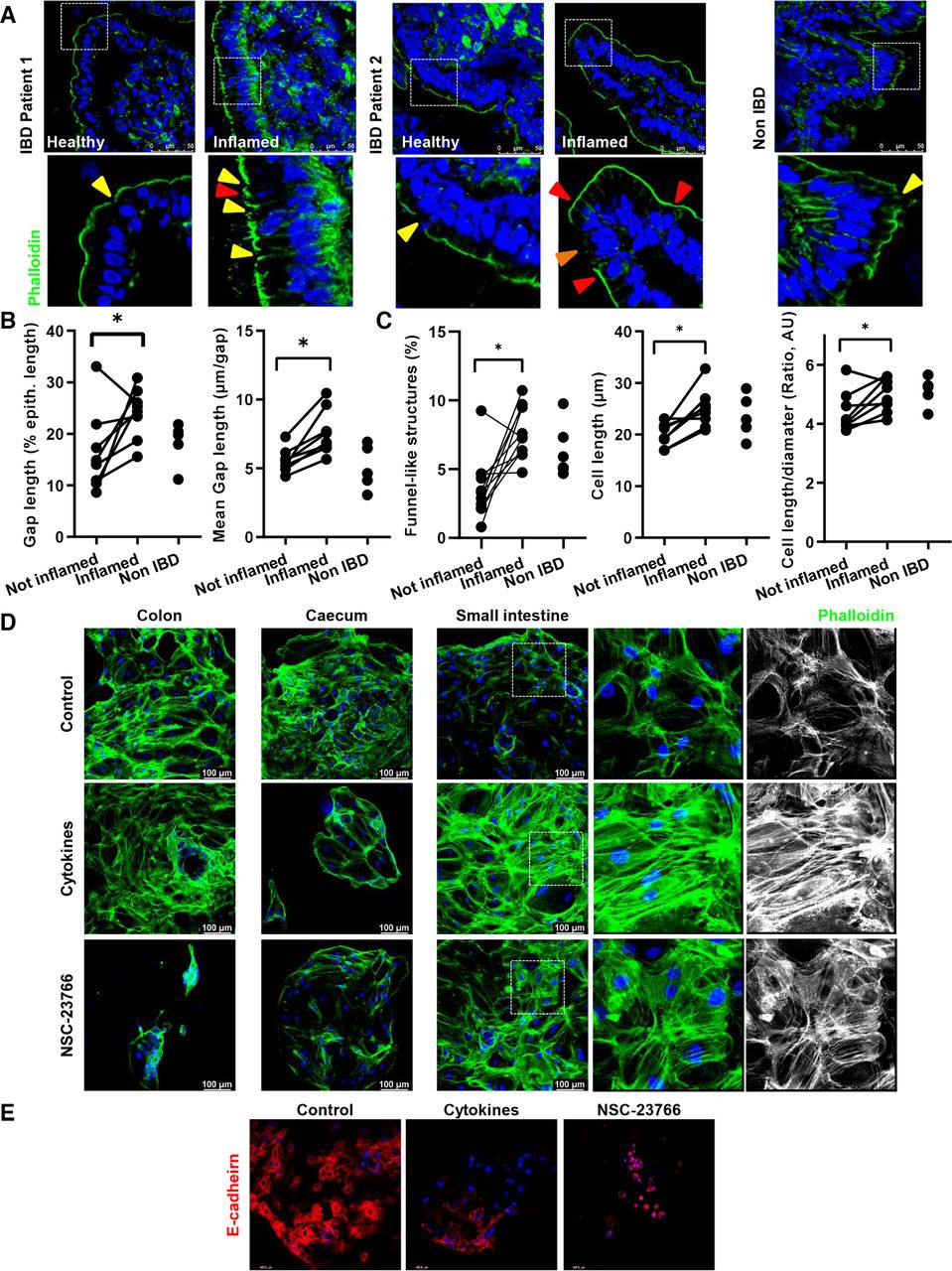
为了提供细胞脱落在人类肠道炎症中的作用的进一步信息,我们分析了IBD患者(活动性疾病和非活动性疾病)和非IBD患者小肠样本中的细胞脱落。我们将屏障功能的改变与Watson评分相关联4 17有细胞脱落或过度拥挤的迹象(图10)。炎症与上皮间隙密度(%间隙/总上皮长度)和间隙长度(平均长度,µm)显著增加(图10 a, B),证实炎症上的上皮渗漏。与此同时,发炎部位的上皮细胞显示大量漏斗状结构(在线补充图2)和累积早期细胞脱落事件的趋势(图10 a, C;在线补充图7);此外,IBD患者IECs的细胞形状改变表现为细胞长度和长度/直径比显著增加(图10 a, C) (在线补充图7)。然后我们想知道这是否与上皮渗漏和/或功能障碍有关。为了验证这一点,我们从另一个人群队列中选择了FABP2血清浓度与e -钙粘蛋白改变(上皮损伤)之间的相关性的患者,46以及漏斗状结构的出现,作为阻止细胞脱落的替代品。有趣的是,尽管样本数量较少,但我们可以观察到FABP2/漏斗状结构之间呈正相关,而E-cadherin表达与FABP2浓度呈负相关(在线补充图7)。总之,这些数据表明了IBD中细胞脱落和上皮渗漏之间的潜在联系。

IBD患者小肠部分细胞脱落和过度拥挤的分析。(A-C)用AlexaFluor488-phalloidin(绿色)染色f -肌动蛋白纤维(n=13,总数;n = 5,控制;n = 8, CD)。(A)代表性图片(上);黄色箭头表示上皮间隙;橙色箭头表示微侵蚀。(B, C)量化。(B)间隙长度(占上皮长度的%),平均间隙长度(µm/间隙)。(C)漏斗状结构(红色三角形)(占总细胞的%)和计算的长度/直径比(AU)。 (D,E) 2D/monolayer organoids generated from human intestinal crypts (three patients), and treated with NSC-23766 (100 µM) or a cytokine cocktail (IL-1β 10 ng/mL, IL-6 10 ng/mL and TNF-α; 20 ng/mL). (D) Phalloidin staining. (E) E-cadherin staining. Paired t-test. *P≤0.050. IBD, inflammatory bowel disease.
接下来,我们的目标是建立rac1介导的细胞骨架重排和上皮渗漏之间的联系。有趣的是,人类的二维类器官47描述了细胞因子鸡尾酒(TNF-α, IL-1β, IL-6)治疗后的形态学变化和TJ改变,模拟炎症肠道中的促炎环境,48但深层肌动蛋白再分配只能在小肠类器官的情况下观察到(图10 d, E) (在线补充图7)。引人注目的是,在使用RAC1抑制剂NSC-23766治疗时,可以观察到类似的上皮改变(图10 d)。在功能水平上,当分别用NSC-23766或GGTI-298处理co2或HT29细胞时,经上皮抗性显著降低,这与肌动蛋白重分布相关(在线补充图7)。因此,我们可以证实RAC1依赖的上皮改变(细胞骨架重排)诱导了类似于炎症肠道中发生的上皮屏障功能的破坏,强调了上皮内在机制的作用,例如由于抑制前烯酰化或RAC1而导致的细胞骨架重排。
我们在人体组织样本和类器官中收集的数据支持IBD中由于上皮细胞骨架/细胞力学缺陷导致的细胞脱落和过度拥挤与慢性肠道炎症之间的联系,证明了在小鼠研究中所确定的机制的临床相关性。
IBD中上皮RAC1通路的改变
对RAC1亚细胞定位的分析表明,在非ibd组织中,上皮表面IECs的根尖膜定位向位于隐窝底部的细胞的细胞质和/或细胞核转移(图11)。这表明RAC1在调节上皮挤压(表面上皮)和细胞增殖(隐窝底)方面可能具有不同的功能。有趣的是,我们可以在IBD患者中观察到RAC1根尖定位的变化,无论是在缓解期还是活动性疾病患者中,炎症表达的降低,都表明RAC1功能障碍(图11)。

人类IBD中RAC1通路的分析(A) RAC1免疫染色(白色),EpCAM(绿色)和Hoechst(蓝色)反染色。代表性图片,显示上皮表面(上)和隐窝(下)的表达和亚细胞定位(n=20,共计;n = 9,控制;n = 11,炎症性肠病)。Wave1 (B)和Wave2 (C)免疫染色(红色)(Wave1, n=13,共;n = 5,控制;n = 8, IBD);(Wave2, n=17, total;n = 6,控制; n=11, IBD).
为了确认rac1依赖通路与维持肠道上皮完整性的潜在相关性,我们分析了WAVE蛋白和rac1下游通路的激活。在体内,特别是WAVE2和WAVE1在回肠组织早期表达显著降低Rac1iec中的删除Rac1我ΔIEC老鼠(在线补充图5),表明RAC1的存在对于WAVE蛋白的表达和/或稳定性是不可或缺的。值得注意的是,在IBD患者的上皮细胞中也可以观察到WAVE蛋白的改变。IBD患者上皮WAVE1表达降低(图11 b)。此外,炎症时上皮表面没有均匀的WAVE2表达,而绒毛/隐窝轴向下和隐窝中没有观察到变化,表明上皮表面特异性的RAC1功能障碍(图11 c)。有趣的是,我们可以从实验性结肠炎的动物中观察到WAVE2在大肠上皮中的表达下降,如右旋糖酐硫酸钠(DSS) (图12)或过继性淋巴细胞转移性结肠炎(图12 b);这与炎症程度相关,主要见于上皮表面。Western blot分析证实,从dss诱导的结肠炎小鼠分离出的IECs中,WAVE2表达下调(图12)。自Wasf1而且Wasf2在DSS或TC小鼠的结肠IECs中,基因表达没有改变,在小肠GGTase或rac1缺陷类器官中也没有改变(在线补充图8),我们假设rac1依赖的Wave蛋白调控是通过蛋白稳定性和/或翻译后机制介导的。由于WAVE蛋白的稳定性是通过蛋白酶组降解来调节的,49然后我们测试了蛋白酶体(MG-132)抑制对WAVE降解的影响以及对细胞脱落改变的影响。在缺乏rac1的小肠类器官中,被阻止的细胞脱落事件的积累被MG132蛋白酶体抑制部分挽救(图13)。同样,在RAC1抑制剂NSC-23766的治疗中,漏斗状结构的积累与人类类器官中WAVE2表达的降低有关(图13 b, C)。这种肌动蛋白的改变仅在MG132处理时得到部分修复,这与蛋白酶体抑制不能修复人体类器官中nsc23766介导的RAC1抑制中的WAVE2表达这一事实相一致(图13 b, C)。综上所述,这些数据表明RAC1功能和下游通路可以促进IECs内的细胞骨架重排,特别是在上皮表面和/或绒毛尖端,以维持IBD患者肠道上皮的完整性,其机制与细胞脱落改变有关。

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
实验性结肠炎RAC1通路分析小鼠实验性结肠炎模型中WAVE1和WAVE2的表达。(A) dss引起的结肠炎。左图为结肠样本免疫染色的代表性图片,右图为分离IECs的WB图像。免疫染色(n=10,共;n = 5,控制;n = 5);WB (n=13,合计;n = 6,控制;n = 7, DSS)。 (B) Adoptive lymphocyte TC. Representative pictures from immunostaining in colon samples (n=14, total; n=6, Control; n=8). Data are expressed as mean±SEM. Unpaired t-test. *P≤0.050. TC, transfer colitis.
{kind=link}

{kind=link}
干扰类器官中的RAC1通路。(A)用蛋白酶体抑制剂MG132(1µM)处理rac1缺陷小肠类器官。Phalloidin染色。三个实验。(B,C)用RAC1抑制剂NSC-23766(100µM)和蛋白酶体抑制剂MG132 (n=3)处理的人体类器官。(B) Phalloidin染色。(C)波2染色。
讨论
在这里,我们证明了细胞骨架重排和细胞脱落紊乱是引发通透性增加的致病现象,最终导致肠道炎症和组织结构的破坏。因此,阻止生理性细胞脱落的上皮渗漏会引发肠道炎症。早期细胞骨架重排对上皮细胞ggtase -戊烯酰化的抑制作用Pggt1b我ΔIEC老鼠34细胞挤压完成受损,导致细胞脱落受阻,细胞过度拥挤,出现“可渗透”细胞(图14)。这种表型主要影响小肠,较小程度上影响结肠。本研究的新颖之处在于,GGTase-deficient和RAC1-deficient上皮细胞的上皮变化仅由细胞内在现象触发,但也会导致肠道炎症。就像tnf诱导的上皮改变一样,50肠组织Pggt1b我ΔIEC而且Rac1我ΔIEC小鼠显示细胞骨架/TJ改变,随后上皮破坏和屏障破坏。缺乏RAC1和ggtase的类器官显示细胞骨架重排和上皮完整性丧失,表明在缺乏外部因素的情况下原发性上皮改变。有趣的是,戊烯酰化抑制或RAC1缺陷上的AJC蛋白重分布反映了人类肠道炎症中的上皮功能障碍,即claudin-1的变化,51claudin-2,52claudin - 826claudin-18,53钙粘蛋白,54β-catenin和ZO-1。55总之,我们的数据强调RAC1作为ggtase靶点有助于上皮完整性和肠道稳态。
{kind=link}

{kind=link}
抑制肠上皮细胞内GGTase1或RAC1诱导肠道炎症的机制图已创建与BioRender.com。
我们的研究揭示了IBD中生理细胞脱落和上皮RAC1通路的改变与肠道炎症之间的潜在相关性。在我们的人类队列中,屏障功能障碍(上皮间隙)的迹象伴随着IBD患者肠道炎症区域漏斗状结构和细长细胞形状的积累,这表明IBD炎症与阻止脱落和细胞过度拥挤之间存在关联。值得注意的是,这也可能与上皮渗漏有关,如血清FABP2水平升高和e -钙粘蛋白改变所示。此外,nsc -23766诱导的RAC1抑制导致细胞骨架重排和TJ分解(人类类器官)和上皮抗性受损(人类上皮细胞系)的事实证实了上皮屏障功能在上皮内在机制上的破坏,如由于抑制丙烯酰化或RAC1而改变细胞力学。值得注意的是,我们对患者材料的分析首次表明,除了病理性细胞脱落外,控制活细胞挤出可能在慢性肠道炎症的发病机制中发挥关键作用,应在IBD的背景下进一步探索。
生理性脱落与病理性脱落的分离与细胞凋亡密切相关。56由于TJs表达调控caspase活性和凋亡细胞死亡,57我们假设细胞骨架重排和TJs重分配可能导致GGTase1B和rac1缺陷上皮细胞中caspase-3切割减少。此外,凋亡和caspase独立细胞死亡之间的转换提示了GGTase-或rac1缺陷上皮细胞中替代细胞死亡途径的潜在激活。在GGTase和/或rac1缺陷小鼠中,我们的发现与细胞死亡激活对肠道损伤的贡献是一致的,但我们的数据表明,这应该被认为是导致进一步上皮损伤和组织破坏的额外机制。
缺乏rac1的肠道类器官的转录组变化与机械刺激的调节反应相关,提出了rac1介导的上皮完整性的机械生物学的关键功能。实际上,缺乏ggtase和缺乏rac1的IEC显示了细胞形状的改变(TEM),而抑制烯丙化与IEC变形能力的变化(RT-FDC)相关。三维牵引力显微镜分析表明上皮细胞骨架变化与ECM力学之间存在相互作用。与WT类器官相比,由Pggt1b我ΔIEC而且Rac1我ΔIEC小鼠表现出向外定向的基质变形,表明力松弛。因此,GGTase和rac1缺陷上皮细胞骨架重排影响IEC和/或ECM机制。此外,细胞骨架动力学也可以直接影响基因的转录。58在我们的RNASeq分析中,几种促进免疫细胞募集的趋化因子出现在rac1缺陷类器官中最受调控的基因中(CXCL-1, CXCL-2和CXCL-10)。总之,rac1依赖的上皮细胞机制成为理解IECs与其细胞外环境之间相互作用以及如何影响上皮-免疫通信的重要上皮-内在机制。上皮细胞机制有助于肠道组织内稳态,应在人类疾病(如IBD)的背景下进一步研究。
肠道疾病严重程度的差异Pggt1b我ΔIEC而且RhoAΔIEC老鼠34提示RHOA与其他戊烯酰化靶标之间存在代偿功能重叠。因此,受损的RAC1功能模拟了烯丙化抑制的上皮改变。一些研究旨在破译RHOA/RAC1/ cdc42依赖的肌动蛋白“钱包”和RAC1介导的细胞爬行对上皮间隙闭合的贡献。31日37考虑到间隙几何形状,负曲率促进rhoa依赖的肌动蛋白环闭合,而正曲率指示rac1介导的细胞爬行。59与我们的数据一致,由于绒毛尖端的凸几何,上皮细胞的挤压将受到RAC1功能的调节。此外,研究表明,小鼠小肠中依赖RAC1的细胞骨架重排控制隐窝/绒毛隔室化,而RAC1的缺失导致肠道结构紊乱。60这些观察结果与我们在体内rac1依赖的上皮完整性和肠道组织结构维持的数据一致。
在绒毛/隐窝轴上,RAC1普遍存在于隐窝底部和瞬时扩增区(transient- amplification area, TAA),而膜/细胞质蛋白则局限于绒毛。有趣的是,在IBD炎症中RAC1的亚细胞转移以及上皮WAVE2表达的下调也仅限于上皮表面,并可能与膜RAC1的特定改变有关。RAC1还可以转位到细胞核,参与细胞周期调节和核细胞骨架特征,61从而控制癌症的发展。62目前尚不清楚戊烯酰化如何分别影响活化的RAC1的膜/核定位,以及在炎症或癌症的过度增生中如何调节上皮屏障。
验证RAC1在肠道稳态中的相关性,Rac蛋白编码基因的改变63 64和RAC1信号65都与IBD有关此外,硫唑嘌呤依赖的Vav/RAC1靶向诱导T细胞凋亡,因此目前被用作IBD患者的治疗方法。66尽管有特定的RAC1抑制剂,67 68其他人69我们的数据假设RAC1激活有助于保护上皮的完整性。因此,调节RAC1功能可以为诊断、预测和/或预防IBD发作的生物标志物的鉴定铺平道路。70并被用于上皮细胞的恢复。71 72然而,IECs和免疫细胞中潜在的相反作用强调了细胞特异性研究对优化Rho GTPases的药理学靶向的相关性。34 73 74
数据可用性声明
所有与研究相关的数据都包含在文章中或作为补充信息上传。
伦理语句
患者发表同意书
伦理批准
本研究为人类受试者,由埃尔兰根-纽伦堡大学(Erlangen, Germany)医学系伦理委员会批准(文献编号440_20 B),受试者在参加研究前知情同意参加。
致谢
目前所做的工作(部分)满足了获得博士学位的要求。nat. for Luz del Carmen Martínez Sánchez, Phuong A Ngo和Rashmita Pradhan。我n some cases, microscopy/Image analysis was performed with the support of the Optical Imaging Centre Erlangen (OICE) and Philipp Tripal. Confocal microscopy was performed on a Leica Stellaris 8 system, funded by Deutsche Forschungsgemeinschaft (DFG, German Research Foundation)—project 441 730 715.
参考文献
脚注
推特@AlastairWatson3
最惠国和自由人民党同样作出了贡献。
贡献者RP、LdCMS、PAN和L-SB进行了大部分实验。RP, TK, VT, LE, IS, CBe, CG和BW为我们的一些实验提供了帮助和支持。DB和BF进行了三维牵引力显微镜测量。DS、MKu、CS和JG进行了RT-FDC实验。MKl、CD、KA进行电镜实验。ST进行了质谱实验。RA提供了人体材料/样品。AJMW和IA阅读了手稿,并为最终版本做出了贡献。MB和CBr提供了鼠标线。MFN和RP设计了研究并撰写了手稿。 RLP is responsible for the overall content as guarantor. All authors approved the final version of the manuscript.
资金本研究由德国研究机构(DFG)资助;批准号(LO-2465/1-1, SPP-1782-LO-2465/2-1, TRR241-A07, TRR-SFB225 (A01),埃尔兰根-纽伦堡大学新兴领域计划)和埃尔兰根-纽伦堡大学医院(ELAN)跨学科临床研究中心(IZKF)。
相互竞争的利益没有宣布。
出处和同行评审不是委托;外部同行评审。
补充材料此内容由作者提供。它没有经过BMJ出版集团有限公司(BMJ)的审查,也可能没有经过同行评审。讨论的任何意见或建议仅是作者的意见或建议,不被BMJ认可。BMJ不承担因对内容的任何依赖而产生的所有责任和责任。如果内容包括任何翻译材料,BMJ不保证翻译的准确性和可靠性(包括但不限于当地法规、临床指南、术语、药品名称和药物剂量),并且对因翻译和改编或其他原因引起的任何错误和/或遗漏不负责。
请求的权限
如果您希望重用本文的任何或全部内容,请使用下面的链接,该链接将带您到版权清除中心的RightsLink服务。您将能够快速获得价格和即时许可,以多种不同的方式重用内容。