条文本

摘要
目标肠道菌群是肥胖和2型糖尿病的关键组成部分,但这种相互作用的机制和代谢物仍不清楚。我们在MetaCardis队列中检测了健康代谢状态和肥胖和2型糖尿病最严重状态下的人类肠道微生物组的功能组成。我们重点研究了B族维生素和B7/B8生物素在调节宿主代谢状态中的作用,因为这些维生素影响微生物功能和宿主代谢和炎症。
设计我们对来自MetaCardis队列的1545名受试者和不同的小鼠实验进行了宏基因组分析,包括无菌和抗生素处理的动物、粪便微生物群转移、减肥手术和在小鼠中补充生物素和益生元。
结果严重肥胖与细菌生物素产生体和转运体的绝对缺乏有关,它们的丰度与宿主代谢和炎症表型相关。我们发现重度肥胖中循环生物素水平低于理想水平,人类脂肪组织中生物素相关基因表达改变。在小鼠中,抗生素引起的肠道菌群缺失或缺失证实了微生物对宿主生物素水平的贡献。减肥手术可以改善人体和小鼠的新陈代谢和炎症,增加细菌生物素的产生,并改善宿主系统生物素。最后,给高脂日粮喂养的小鼠补充低聚果糖和生物素,不仅可以改善微生物组的多样性,还可以提高细菌产生生物素和B族维生素的潜力,同时限制体重增加和血糖恶化。
结论结合生物素和益生元补充的策略可以帮助防止严重肥胖的代谢状态恶化。
试验注册号码NCT02059538.
- 肥胖
- 微量元素
- 营养
- 肠道细菌
- 糖尿病
数据可用性声明
数据可在一个公共的、开放访问的存储库中获得。根据合理的要求提供数据。metacardishttps://www.ebi.ac.uk/ena/browser/view/PRJEB41311,https://www.ebi.ac.uk/ena/browser/view/PRJEB38742,https://www.ebi.ac.uk/ena/browser/view/PRJEB37249老鼠实验;16 s数据,https://www.ebi.ac.uk/ena/browser/view/PRJEB42967老鼠实验;纳米孔的数据,https://www.ebi.ac.uk/ena/browser/view/PRJEB42966,代码论文分析,https://git.ummisco.fr/ebelda/metatransitionbiotingut.git.
这是一篇开放获取的文章,按照创作共用署名非商业性(CC BY-NC 4.0)许可发布,该许可允许其他人以非商业性的方式发布、混编、改编、构建本作品,并以不同的条款授权他们的衍生作品,前提是原创作品被正确引用,给予适当的荣誉,任何更改都被注明,且使用是非商业性的。看到的:http://creativecommons.org/licenses/by-nc/4.0/.
来自Altmetric.com的统计
本研究的意义
关于这个问题,我们已经知道了什么?
肠道菌群通过产生许多代谢物,包括细菌来源的B族维生素,影响代谢健康。细菌产生B族维生素的受损会影响微生物群落功能和宿主代谢和炎症。然而,这些方面还没有在严重肥胖和失调的背景下进行探索。
啮齿动物和临床研究表明,在肥胖和代谢性疾病中,血清和组织生物素状态发生改变。
肥胖性饮食已知会诱导严重的生物失调,但尚不清楚它是否也会导致细菌代谢的伴随改变和宿主生物素利用率的降低。
本研究的意义
新的发现是什么?
MetaCardis是一群患有严重肥胖和代谢改变的大型欧洲人群,对该人群的宏基因组分析表明,在这些条件下观察到的微生物细胞负荷的损失与人类生物素生物合成和运输的细菌潜力的改变有关。
细菌生物素状态的改变也与严重肥胖的代谢和炎症表型有关。
高脂饮食诱导的肥胖导致小鼠体内微生物和宿主生物素状态的改变。
无菌、抗生素处理的SPF小鼠和人对小鼠肠道菌群转移实验证明了肠道菌群对宿主血清生物素状态的贡献。
减肥手术引起的体重减轻改善了微生物生物素代谢和宿主生物素状态。
口服生物素和益生元制剂可改善小鼠体重、葡萄糖代谢和生物素微生物状态。
在可预见的未来,它会对临床实践产生怎样的影响?
提高在代谢性疾病的临床治疗中同时考虑肠道菌群和宿主生物素状态的意识。
通过以肠道为中心的治疗(如益生元)和B族维生素的可用性(包括生物素)来同时管理肠道菌群失调,似乎对防止肥胖过渡到更严重的代谢状态很有意义。
简介
重度肥胖,定义为体重指数(BMI)≥35 kg/m2,从1975年到2014年,全球男性增加了11倍(2.3%),女性增加了3倍(5.0%)。1并因其相关的心脏代谢并发症而承担沉重的负担。2组织改变(如代谢改变和炎症增加)和全身低级别炎症是人们熟知的将严重肥胖与2型糖尿病(T2D)联系起来的因素。越来越多的证据表明,肠道菌群是导致这些现象的一个因素。从超重到重度肥胖,肠道微生物基因丰富度普遍降低3.随着代谢并发症的严重程度而增加。4我们最近发现,在欧洲MetaCardis队列的一个子集中,严重肥胖中拟杆菌2 (bact 2)肠道型的富集与炎症标志物相关。5因此,肥胖和葡萄糖代谢恶化具有宏基因组物种特征,4 - 6它的功能潜力还需要更好的解读。
肠道菌群可以通过产生许多代谢物直接或间接地影响代谢健康,7包括来自细菌的维生素B这表明,细菌产生B族维生素的能力受损会影响两种微生物群落功能8以及宿主代谢和炎症9日10而维生素B7/生物素在重度肥胖中的研究却很少。11
在微生物区系中,生物素代谢和循环的内稳态是细菌正常生长和功能的关键。事实上,肠道细菌形成了复杂的生态系统,其中营养网络具有互补的代谢特征,包括维生素交叉喂养;不能合成维生素的细菌生长所必需的;例如,营养不良的生物素转运体。12从外部来源使用维生素在能量上比合成维生素更有利。实际上,产生一个生物素分子(如原营养真核生物)需要超过6种酶和7种ATP等价物。13有趣的是,生物素的生物合成被描述为富含拟杆菌的微生物组组成的特征。14这与原核生物中原营养生物素生物合成的拟杆菌系的富集是一致的,在b族维生素代谢的不同基因组调查中有描述。8日15此外,在包括肥胖和T2D在内的改变的健康状况中,也描述了微生物组生物素生物合成潜力丰度的增加。16然而,这些分析仅仅基于相对丰度宏基因组谱。然而,正如在严重肥胖中观察到的那样,富含拟杆菌的微生物群(通常是从相对丰度剖面定义的)与低微生物细胞密度密切相关,5 17这对宏基因组特征的量化以及随后与临床表型的关联有着强烈而显著的影响。17我们可以假设,像生物素生物合成潜力等宏基因组特征的丰度,在拟杆菌属中高度富集,可能会受到生态系统的细胞密度的严重影响。因此,通过考虑微生物细胞密度来评估微生物组生物素生产和转运潜力的绝对定量,可以揭示重度肥胖中微生物生物素的状态。
目前尚不清楚生物素细菌代谢是否在严重肥胖中发生改变,以及肠道菌群失调是否会影响宿主的生物素状态。生物素是宿主生理功能的关键,包括碳水化合物和脂质代谢。18生物素是羧化酶的辅助因子,羧化酶在脂肪酸合成和线粒体氧化中起关键作用,包括在人体脂肪组织中。19在中度肥胖和T2D患者中,血清生物素与对照组相比有所下降,20.独立于宿主遗传背景。20.致肥性饮食,已知可诱导严重的生物失调,是否也导致伴随的细菌代谢改变和生物素对宿主的可利用性降低尚不清楚。
补充生物素可以改善T2D受试者和啮齿动物的糖代谢,18研讨会并影响脂肪组织的新陈代谢。口服生物素补充小鼠模型显示代谢改善和改善一些生物素依赖性羧化酶的组织表达。24日25日然而,在西化饮食引起的肥胖或严重肥胖人群的背景下,它还有待深入研究。
考虑到微生物细胞的失调和损失,我们的目的是通过横断面分析1545名处于代谢恶化不同阶段的肥胖受试者的粪便微生物群,来表征严重肥胖中的细菌生物素代谢潜力,并研究其对宿主代谢状态的重要性。5通过一系列小鼠实验,包括纠正生物素状态对肠道菌群失调的影响,我们进一步深入了解了细菌生物素和宿主的交叉对话。
结果
定量宏基因组学关联揭示了重度肥胖中生物素合成和转运潜力的改变
我们比较重度肥胖患者(SOB, BMI≥35 kg/m²;n=608)为超重或I级肥胖(MOB, BMI 25-34.9 kg/m²;n=299)和苗条的非肥胖者(NOB, BMI <25 kg/m²;n = 638)。在这些BMI组中,我们还进一步调查了葡萄糖代谢改变严重程度增加的影响(代谢健康组(MH;n=458),代谢不健康组(MUH;n=430)和T2D组(T2D;n = 657)) (在线补充讨论有关队列描述,请参见在线补充表S1-S4).
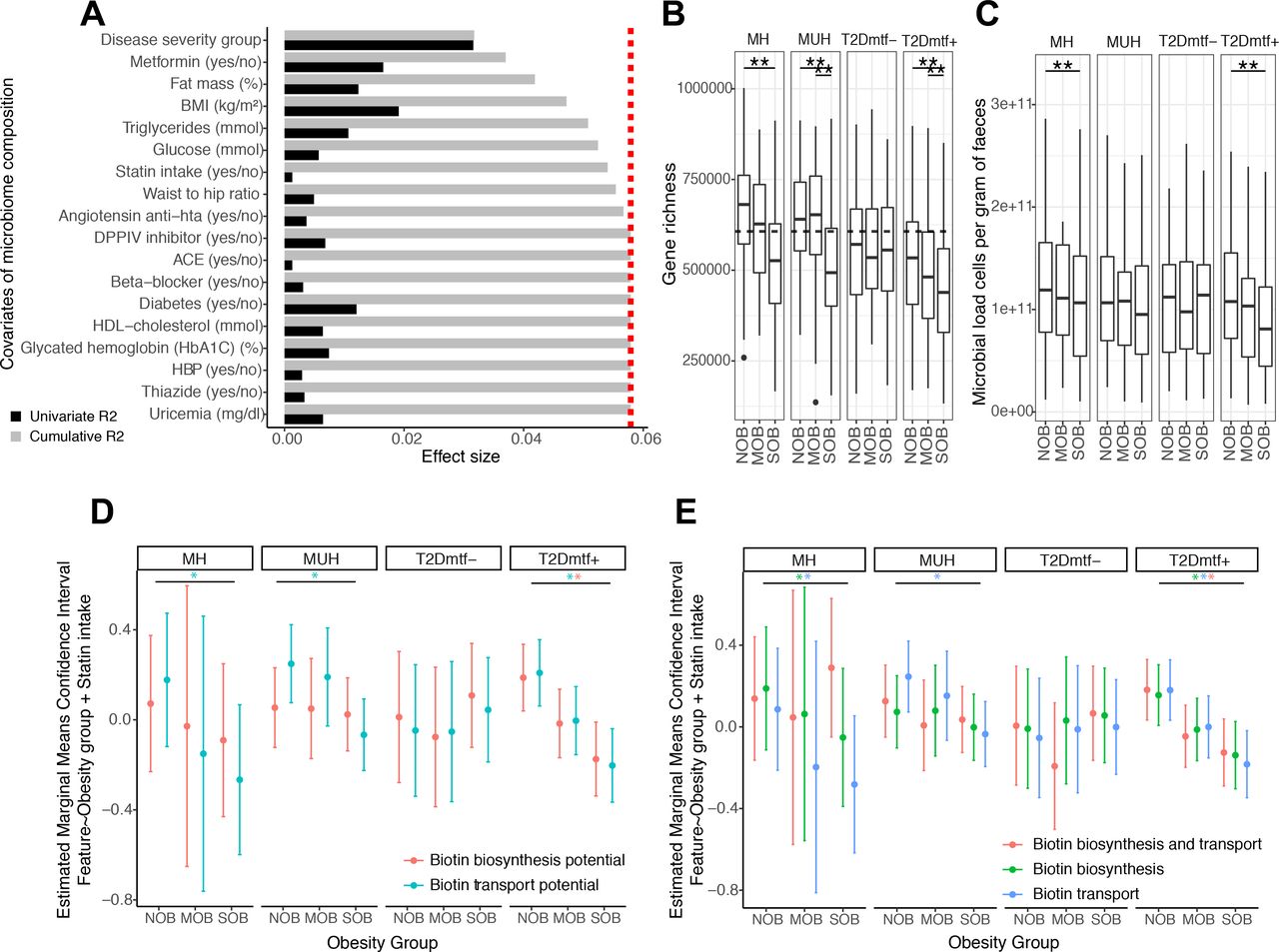
这种按疾病严重程度分层(BMI和葡萄糖代谢恶化)是解释微生物组组成的最高非冗余比例的变量(n=1545,基于逐步距离的冗余分析(dbRDA)与mOTU丰度数据属级相对丰度曲线,R2=3.17,错误发现率(FDR)=1.0×10−4),其次为BMI (dbRDA, R2=0.53, FDR=1.0×10−4),二甲双胍摄入量(dbRDA, R2=0.52, FDR=1.0×10−4),脂肪量百分比(dbRDA, R2=0.48, FDR=1.0×10−4),血清甘油三酯(dbRDA, R2=0.36, FDR=7.0×10−4)和血糖(dbRDA, R2=0.16, FDR=1.82×10−2) (图1一个).他汀类药物也被纳入对微生物组变化无冗余影响的变量(dbRDA, R2=0.16, FDR=2.1×10)−2,图1一个).5

代谢健康组中与肥胖严重程度相关的功能特征:细菌细胞负荷的影响(A)解释MetaCardis队列子集中微生物组组成变化的主要变量(基于距离的冗余分析,dbRDA;属级的布雷-柯蒂斯不相似度),要么独立(单变量效应大小为黑色),要么在多元模型中(累积效应大小为灰色)。对多元模型的重要非冗余贡献的截止点用红线表示。(B)按代谢健康状况分层的肥胖组间基因丰富度分布。(Kruskal- Wallis检验对照的**P值<0.05,FDR检验配对对照的Wilcoxon秩和检验对照的招募国和年龄)。虚线表示根据健康德国人群(n=91)中基因丰富度的中位数(n=91)将个体分层为高与低基因数(HGC/LGC)的阈值,该人群表现出基因丰富度双模态。(C)按代谢健康状况分层的肥胖组微生物细胞计数分布。(Kruskal-Wallis检验对照的**P值<0.05,FDR配对Wilcoxon秩和检验对照的招聘国<0.05)。(D)经他汀类药物摄入调整并按代谢健康状况分层的肥胖组微生物组生物素生物合成和消耗潜力的对数转换绝对丰度的估计边际均值和置信区间。 (E) Estimated marginal means and confidence intervals of log-transformed absolute abundances of biotin producers (eg, prokaryotic organisms harboring all biotin biosynthesis genes from pimelate precursor and no biotin biosynthesis transport genes), biotin transporters (prokaryotic organisms with no biotin biosynthesis genes) and biotin producers and transporters (prokaryotic organisms with all biotin biosynthesis genes from pimelate and biotin transport genes) across obesity groups adjusted by statin intake and stratified by the metabolic health status (*FDR<0.05 on linear regression models of feature abundance by obesity status adjusted by statin intake, P-adj<0.05 on pairwise Tukey tests between obesity states). BMI, body mass index; HbA1c, haemoglobin A1c; HDL, high-density lipoprotein; MH, metabolically healthy; MUH, metabolically unhealthy; T2D, type 2 diabetes.
除未接受二甲双胍治疗的T2D个体外,严重肥胖的特征是代谢组微生物基因丰富度显著降低(图1 b;FDR<0.05)。与其他组相比,该组患者T2D用药明显不足(Χ2P = 7.57×10−3;在线补充图S1).在服用二甲双胍治疗的MH和T2D个体中,严重肥胖的特征是微生物细胞密度显著降低(图1 c;FDR<0.05)。
我们利用定量微生物图谱(QMP)估算了微生物组生物素生物合成和消耗潜力的绝对定量。QMP评价来自于人肠道微生物组综合基因目录(IGC)的基因丰度26并从该种群的微生物细胞密度(见方法)。除未接受二甲双胍治疗的T2D个体外,代谢组中生物素转运潜能的QMP随重度肥胖显著降低(图1 d;FDR<0.05)。同样,在MH组中,生物素生产潜力的QMP随着肥胖严重程度的增加而下降,这种下降在接受二甲双胍治疗的T2D个体中具有统计学意义(图1 d;FDR<0.05)。
当使用原核生物中b族维生素生物合成和运输表型的注释来估计微生物组的生产和消耗潜力时,我们证实了这一观察结果。15在服用二甲双胍组的MH个体和T2D个体中,随着肥胖严重程度的增加,生物素营养不良体(如无生物素生物合成基因的细菌)和严格的生物素原营养体(如有来自于聚氰胺的生物素生物合成基因但无生物素运输基因的细菌)的总绝对丰度降低。图1 eNOB组与SOB组相比,FDR<0.05),在能够产生和消耗生物素的细菌群中观察到类似的关联。
我们比较了基于肠道微生物组基因的生物素生产和消费潜力的绝对定量与基于生物素生物合成和运输基因含量的不同细菌亚群的绝对丰度。15生物素合成的绝对潜力与具有完整生物素合成途径且不参与生物素转运的细菌种类的丰度密切相关(在线补充图S2A;P1组,r=0.909, p<0.05)。该类群主要由变形菌门和拟杆菌门(在线补充图S2B:分别占296种生物的49.32%和43.58%)。相比之下,生物素的绝对消耗潜力与没有生物素生物合成基因和携带来自能量耦合因子运输系统的生物素转运基因BioY的细菌种的丰度密切相关27(在线补充图S2A);r = 0.90, p < 0.05)。这一类群由厚壁菌门(在线补充图S2A:该类群618个生物中的77.93%)。最后,这些生物素营养不良和原营养物质在肥胖人群中的丰度分布可以扩展到其他B族维生素(在线补充图S3).
这一分析表明,在代谢疾病恶化的状态下,微生物细胞负荷的损失与B族维生素(包括生物素)的合成和运输潜力的改变有关。
微生物生物素功能模块与严重肥胖的代谢改变和炎症有关
然后我们假设细菌生物素代谢的潜在功能改变可能与宿主的代谢和炎症表型有关。我们针对53个临床和营养变量,对与生物素转运和生物合成潜力相关的宏基因组特征的log10转化QMP丰度进行线性回归分析(例如,IGC基因绝对丰度,基于参考基因组的生物素产生和转运体群体)。来自脂肪酸中间体(P1组)和来自酰美酸前体(P*, P2组)的生物素营养不良体(A组)和严格的生物素从头生成体(P*, P2组)的绝对丰度与肥胖变量(BMI、脂肪量(%)、腰围、内脏脂肪评分)、糖耐量和代谢相关变量(空腹血糖、糖化血红蛋白(HbA1c)、C肽、甘油三酯和胰岛素抵抗和敏感性替代品)呈负相关,以及炎症状态(C反应蛋白(CRP)、白细胞介素6 (IL-6)、白细胞总数、中性粒细胞和单核细胞计数)(图2,标准化beta系数<0,FDR <0.05)。生物素运输和生产的绝对丰度与微生物基因丰富度、胰岛素敏感性替代物(HOMA-S)和胆固醇相关变量(包括高密度脂蛋白(HDL)-胆固醇(图2,标准化beta系数>0,FDR<0.05)。从IGC基因丰度和参考基因组(图2).这一关联分析支持严重肥胖中宿主代谢、炎症状态和细菌生物素代谢紊乱的内稳态之间的相互作用。

MetaCardis亚队列中微生物组生物素状态与宿主代谢和炎症标志物之间的关系。热图显示生物素生产和运输相关宏基因组特征的log-10转换QMP丰度谱与临床和生活方式因素之间的调整关联。y轴为自变量,x轴上的变量为因变量(n=1545个个体)。这些模型根据招聘国家和年龄进行了调整。(* P值< 0.05;* *罗斯福< 0.05。没有发现与FDR<0.05相关的临床和生活方式变量不包括在热图中)。色调对应于调整后的线性回归模型中标准化beta系数表示的效应大小。根据Rodionov中定义的命名法定义生物合成和运输基因组组等.15简单地说,这包括3组严格的生物素产生者(P1, P2, P*组),它们拥有所有4个共同的基因,用于生物素生物合成的不同途径变体,从聚酸酯(P2)或聚酸酯- acp (P1, P*)。包括8组严格生物素营养不良菌A&S/A组;不能产生生物素的微生物45,具有(A&S组)或不具有(A组)参与生物素转运的基因的微生物),在4个生物素合成核心基因(最多含有1到3个生物素合成基因)和4组同样含有生物素转运编码基因的生物素产生基因(P&S组)中存在不同程度的不完全性。ALT,丙氨酸转氨酶;AST、天冬氨酸转氨酶;BMI,身体质量指数;CXCL5, C-X-C基序趋化因子配体5;GGT,γ谷氨酰转移酶;高密度脂蛋白,高密度脂蛋白。低密度脂蛋白、低密度脂蛋白。
重度肥胖中宿主生物素的改变
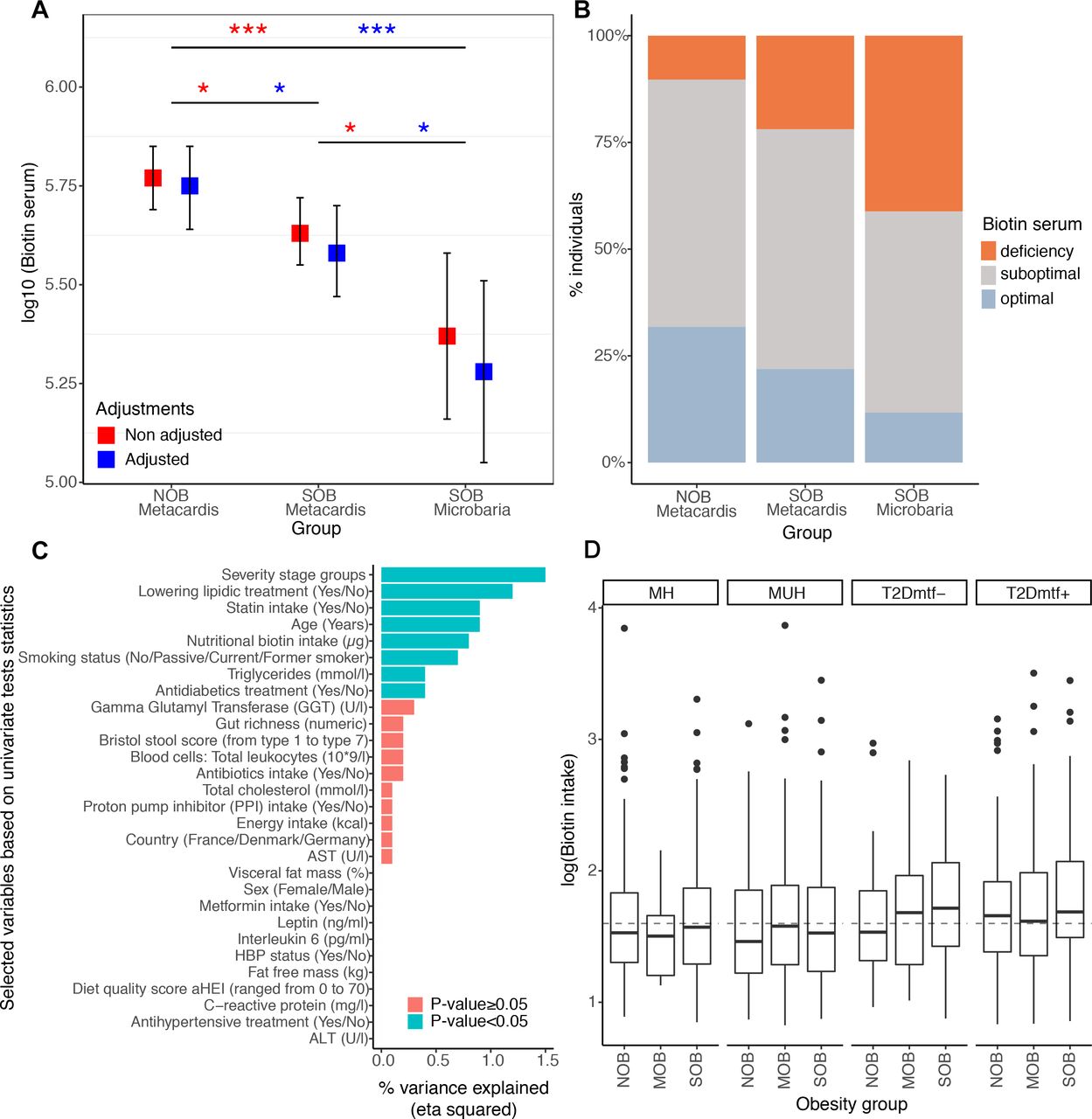
我们研究了严重肥胖中细菌生物素代谢的潜在干扰是否也转化为宿主全身生物素水平。我们对来自独立的Microbaria减肥研究的212名基因丰富度低或高的瘦弱和SOB受试者和17名更严重肥胖受试者的血清生物素进行了量化4(MetaCardis瘦组平均BMI±SD=24.7±3.1,MetaCardis SOB组平均BMI±SD= 41.2±4.9,Microbaria SOB组平均BMI±5.1)。我们发现SOB组的血清生物素有中度但显著的下降(图3一.罗斯福= 3.0×10−2),减少率较高(图3一,罗斯福= 1.12×10−3).我们还使用推荐和发表的阈值评估生物素缺乏。28我们发现,78.09%的MetaCardis SOB组(88.24%的Microbaria组)的血清生物素水平低于最佳水平(血清生物素:200 - 400 ng/L)或缺乏水平(血清生物素:<200 ng/L),而只有19.65%的瘦肉受试者低于该水平(图3 b).我们观察到尿中3-羟基异戊酸的排泄增加,这是早期的标志29生物素缺乏症;符合宿主生物素缺乏。该指标与BMI呈正相关(n=1545), (R2=5.6×10−2, p = 7.9×10−14)或没有(R2=3.0×10−3, p = 3.6×10−2)糖尿病状态、二甲双胍、他汀类和生物素摄入量的调整(线性回归;数据未显示)。在与其他28个临床和营养变量的多变量模型中,疾病严重程度组解释了尿中3-羟戊酸浓度的最高方差分数(图3 c;埃塔的平方= 1.5×10−2;p = 1.2×10−2).然而,在BMI范围内发现的系统性生物素浓度差异不太可能用生物素摄入量的差异来解释。事实上,生物素的摄入来自无处不在的食物来源(例如,主要是肉、鱼、家禽、鸡蛋、乳制品和蔬菜)30.而在肥胖和T2D阶段则略高)(图3 d).

MetaCardis亚队列中肥胖组的全身和营养生物素概况。(A) MetaCardis亚队列中212例肥胖组(n=107 (NOB), n=105 (SOB))和Microbaria研究中17例重度肥胖组之间生物素血清水平的差异(*P值<0.05;* * * P值< 0.001)。非调整和调整(糖尿病状态、二甲双胍、他汀类和生物素的摄入)广义线性模型和lmeans函数的差异显著,并采用Benjamini-Hochberg法进行多次比较的p值调整。生物素血清进行log10转化,使生物素变量(NOB vs. SOB (MetaCardis和Microbaria)) Cohen’s D效应值为0.91。NOB vs. SOB MetaCardis Cohen的效应量D =0.18)。(B)肥胖组间生物素缺乏情况按以下阈值分布:28不足(< 200ng /l)、亚理想水平(200- 400ng /l)、理想水平(> 400ng /l)。Chi-2检验有显著差异(p值=1.0 0x10-2)。(C)临床协变量与尿代谢物3-羟基异戊酸定义的生物素状态之间的关系。横条对应于3-羟基异戊酸的方差由每个临床协变量解释(由来自多元ANCOVA模型的eta平方统计量测量,n=1545)。统计显著性是指一个包含所有变量的全局模型。ALT:丙氨酸转氨酶,AST:天冬氨酸转氨酶,GGT: γ -谷氨酰基转移酶,HBP:高血压。(D)按代谢健康状况分层的肥胖组(n=284 (NOB- mh), n=130 (NOB- MUH), n=51 (NOB-T2Dmtf-), n=173 (NOB-T2Dmtf+), n=13 (mobb - mh), n=81 (mobb -MUH), n=41 (mobb -T2Dmtf-), n=164 (mobb -T2Dmtf+), n=161 (SOB- mh), n=219 (SOB-MUH), n=85 (SOB T2Dmtf-), n=143 (SOB-T2Dmtf+))的log10转化营养生物素摄入量(μg/day)的差异。各组间生物素摄入量无显著差异(FDR>0.05;按国家或他汀类药物摄入量控制的非参数成对单变量试验)。虚线表示欧洲食品安全局(European Food Safety Authority)推荐的每日生物素摄入量(40μg/天)。50;MH,新陈代谢健康;mu,新陈代谢不健康;T2D, 2型糖尿病。
我们下一步假设生物素代谢可能在依赖生物素的宿主组织中发生改变,如脂肪组织。生物素与许多羧化酶共价连接,这种结合由线粒体和细胞质中的全羧化酶合成酶(HLCS)催化,这对组织代谢很重要。19利用之前发布的严重肥胖个体的数据集,31我们观察到BMI与皮下脂肪组织生物素依赖性羧化酶的基因表达呈负相关,如乙酰辅酶a羧化酶(ACACA)、丙酰羧化酶(PCCA、PCCB)、3-甲基crotonyl-辅酶a羧化酶(MCC2)、丙酮酸羧化酶(PC)或HLCS20.(qPCR-数据,Spearman相关性在−0.42和−0.70之间,所有p<0.05;n = 24;在线补充图4).这些基因的表达与炎症基因的表达呈负相关,如趋化因子cc -趋化因子配体2 (CCL2)、IL-18结合蛋白、金属肽酶9、分泌型磷蛋白1和肿瘤坏死因子(TNF)受体超家族成员1A (在线补充图4).总的来说,在严重肥胖中观察到系统性和脂肪组织生物素状态的相对不足。
致肥性饮食改变了微生物区系和宿主生物素状态
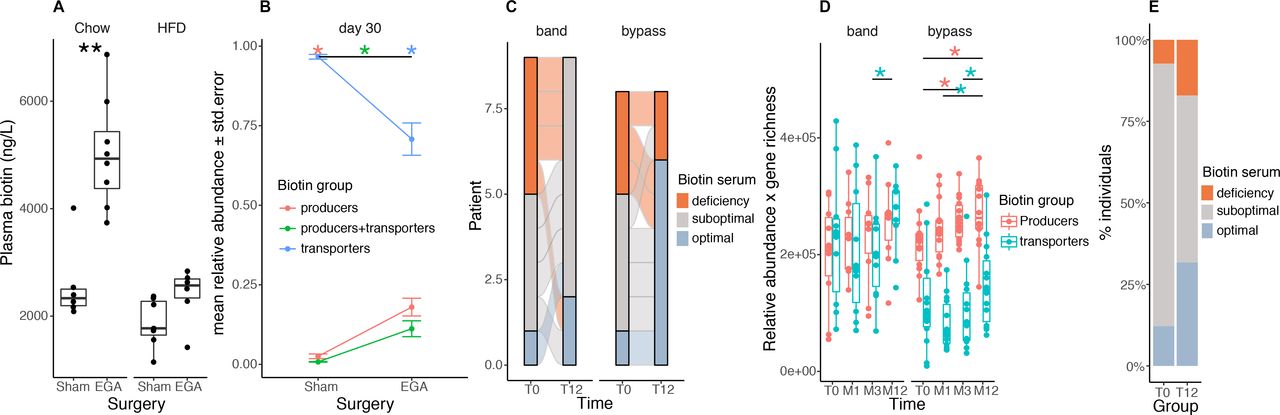
接下来,我们评估了高脂肪饮食(HFD)带来的致肥性条件是否会诱导小鼠体内微生物生物素代谢和宿主生物素利用率的改变。高热量饮食(60%千卡由脂质提供)诱导的肥胖与血浆生物素水平显著降低相关,尽管高热量饮食的生物素摄入量是饲粮动物的两倍(分别为0.65 vs 0.26 μ g /天)。饲喂hfd 4周后,小鼠的血浆生物素水平较饲料喂养的动物有所下降(图4一,左,周与HFD: p=7.32×10−2),在喂养hfd 13周后达到显著水平(图4一右,周星驰vs HFD: p=3.72×10−3).与人类一样,我们发现这些小鼠附睾脂肪组织中生物素羧化酶的表达显著减少(在线补充图5A).在微生物组水平上,经过4周的HFD后,生物素产生体(有生物素生物合成潜能,但没有生物素运输基因)、生物素产生体和转运体(有生物素生物合成和转运基因)的丰度显著下降,同时生物素转运体(没有生物素生物合成基因的细菌)的丰度增加(图4 b;罗斯福= 4.66×10−4, 9.32×103, 4.66×104,分别;Wilcoxon秩和(WRS)检验)。HFD喂养13周后也有相似的结果(图4 b;各组FDR均<0.05)。重要的是,考虑到细菌的总丰度(用qPCR测量),作为细菌负荷的代理,确认了生物素生产者潜力在HFD喂养13周后下降(在线补充图5E,富兰克林·德兰诺·罗斯福< 0.05)。The increase in the abundance of biotin transporters was explained by the large increase ofLactococcus lactisHFD加料后(在线补充图5B),在不同的研究中,它被描述为一种食品污染物。32然而,当l . lactis被排除在微生物组概况之外(在线补充图6).

hfd诱导的小鼠肥胖导致血清生物素水平的减少以及细菌生物素生产谱系的减少。(A)年龄匹配的周食和hfd喂养的C57BL6/J小鼠在4周(左图)和13周(右图)后的血浆生物素浓度(**P值<0.01;第35天和第90天Chow n=7,第35天HFD n=5,第90天n=8, Wilcoxon秩和检验)(B)基线时(第1天),这些小鼠中生物素产生者(具有所有生物素生物合成基因,无生物素转运基因的细菌)、生物素转运体(无生物素生物合成基因的细菌)和生物素产生者+转运体(含有生物素生物合成和转运基因的细菌)的相对丰度谱,第35天和第90天(*P值和FDR<0.05)。(C)无菌(GF)和常规饲养(con - r) C57BL6/J小鼠血清生物素浓度(*P值<0.05,C57BL6/J GF n=7, con - r n=5;Wilcoxon rank-sum测试)。(D)分别用(n=7)和(n=8)大光谱抗生素(100mg/kg万古霉素和200mg /kg氨苄西林、新霉素和甲硝唑)在水中稀释14天,用qPCR法测定小鼠血浆生物素浓度和(E)细菌16S rRNA总基因载量(*P值<0.05);Wilcoxon rank-sum测试)。(F)饮食、表型和生物素生产和运输丰度之间的多变量线性回归模型得出的beta系数,从g组和h组小鼠的粪便转移实验中得出的16S数据和具有所有协变量(*P值<0.05)的同一全局模型中的血清生物素。(g)用来自MetaCardis亚队列的4名受试者的粪便浆定植的瑞士韦氏小鼠的血清生物素水平(2 NOB;2呜咽)。 Mice were colonized for 28 days and were fed either chow (NOB, n=16; SOB, n=12) or western diet (NOB, n=17; SOB, n=17) (*P value and FDR<0.05; ***P value<0.001 and FDR<0.05; Wilcoxon rank-sum test). (h) Abundance of biotin production module inferred from PICRUSt functional profiles of 16S rRNA gene amplicon data of mice from panel F (**P value<0.05; Wilcoxon rank-sum test). CONV-R, conventionally raised; GF, germ-free; HFD, high-fat diet; ns, not significant; WD, Western diet.
肠道菌群对宿主血清生物素有贡献
我们评估了肠道菌群是否有助于生物素循环水平。首先,日粮喂养的无菌(GF)小鼠血清生物素显著低于常规饲养(convl - r) C57BL/ 6j小鼠(图4 c;C57BL / 6 j p = 1.01×10−2).其次,我们同时使用了多种广谱抗生素33给C57BL/6 J小鼠饲喂标准周粮后,血浆生物素显著降低(图4 d;p = 3.11×10−4).我们通过qPCR证实抗生素处理后粪便细菌载量降低(图4 e;p = 2.0×10−4).第三,为了确定人类肠道菌群的不同配置是否会对生物素水平产生不同的影响,我们在瑞士韦伯斯特GF小鼠上定植了两名严重肥胖(SOB)和两名瘦弱(NOB)受试者的粪便,与SOB受试者相比,这两名受试者的粪便中含有更多的细菌生物素产生者。为了考虑饮食的影响,我们给老鼠喂食chow饮食或西式饮食(从脂肪中摄取热量40%)。在与饮食、供体表型和生物素生物合成和运输模块丰度的线性回归分析中(来自16S rRNA基因测序数据),我们发现饮食对生物素血浆水平的影响最大(图4 f;p = 3.2×10−4).然而,在两组用杂粮喂养的小鼠中,我们观察到接受SOB患者肠道菌群的小鼠的生物素血浆水平显著下降(图4 g;p = 1.0×10−2).我们重现了在肥胖性饮食(WD)下小鼠血清生物素水平的下降,这种下降在接受NOB患者肠道菌群(图4 g;p = 4.1×10−4).对于相同的生物素膳食摄入量,肠道菌群的组成影响生物素循环水平。定植小鼠粪便样本的16S测序证实,在供体粪便微生物群中观察到的生物素产生潜能的差异,已转移到饲粮小鼠(图4 h;p值= 4.39×10−3),但对喂食wd的小鼠没有影响。这些结果表明,这种饮食对从供体到小鼠的微生物组功能潜能的转移有显著影响,但我们不能排除肥胖本身的影响。
减肥手术后肠道菌群和宿主生物素状态改善
我们探索了严重肥胖管理背景下的肠道菌群生物素代谢。我们专注于减肥手术(BS),除了改善新陈代谢和炎症外,已知可以诱导肠道菌群变化。4 34我们对饲喂饲料或高脂饮食的C57BL/ 6j小鼠进行了肠胃吻合术(EGA)。35我们观察到,与假手术的动物相比,在bs后3个月(在杂粮喂养的动物中)和1个月(在hfd喂养的动物中)血浆生物素显著增加(图5一个,左:chow, p=1.33×10−3;右:HFD, p = 7.34×10−2).EGA后,饲粮中血浆生物素的增加高于hfd喂养的动物,进一步表明饲粮在调节生物素水平方面的重要贡献。我们对hfd喂养的手术动物的肠道菌群进行了测序,观察到携带或不携带生物素转运基因的生物素生产者的相对丰度显著增加(图5 b,罗斯福= 5.83×10−4在两组)。在考虑到细菌总量(数据未显示)的情况下,也进行了类似的观察。重要的是,这些观察结果在人类中得到了重复,Roux-en-Y胃旁路术(RYGB)术后1年生物素状态最佳的患者数量(来自Microbaria研究)与基线水平相比有所改善(图5 c, p = 2.42×10−2).我们发现,在BS后,产生生物素的细菌显著增加,与宿主生物素状态的改善平行(图5 d;p = 4.94×10−2T0-M3, p = 3.55×10−2T0-M12)。我们在一个独立的患者组中复制了血清生物素的增加。图5 e, p值= 2.0×10−2).与动物模型不同,BS后的患者补充了包括生物素在内的维生素组合(150 μ g/天)。我们可以推测,这种口服补充有助于提高细菌生物素产生的潜力,并改善宿主生物素状态至少在产后的第一年;尽管BS与维生素吸收不良有关。36

小鼠和人实验中减肥手术后生物素的代谢。(A)低脂或高脂饮食(HFD)喂养的C57BL/6J小鼠的血浆生物素浓度,并进行假干预(sham)或减肥手术(肠胃吻合,EGA)。35HFD组于术后1个月采血,Chow组于术后3个月采血(**P值<0.01 Wilcoxon秩和检验;Chow-Sham n=6, Chow-EGA n=8, hhd - sham n=7, hhd - ega n=6)。(B) HFD组假手术小鼠和EGA小鼠术后30天生物素产生体(具有所有生物素生物合成基因但无生物素转运基因的细菌)、生物素转运体(无基因参与生物素生物合成的细菌)和生物素产生体+转运体(携带生物素生物合成和转运基因的细菌)的平均丰度(*FDR<0.05,配对Wilcoxon秩和检验)。(C) Microbaria研究中17例患者在基线至12月间生物素缺乏组的分布情况,按手术组分层(n=9,胃束带;n=8, Roux-en-Y胃旁路术)根据以下阈值:28不足(< 200ng /l)、亚理想水平(200- 400ng /l)、理想水平(> 400ng /l)。P值=2.4 × 10-2(旁路),P值=1.1x10-1(频带);费雪的测试。(D) Microbaria研究中按手术类型分层的24例个体(可调节胃束带,n=10;Roux-en-Y胃,n=14),在减肥手术后基线、1、3和12个月的宏基因组学数据(*P值<0.05;Wilcoxon符号秩检验)。(E) BARICAN队列中搭桥术后基线(T0)和12个月(T12)生物素缺乏组的分布(n=41;P值=2.0x10-2, Chi2检验)。EGA entero-gastro吻合;HFD、高脂肪饮食。
结合益生元和生物素改善微生物区系生物素和代谢状态
这些结果促使我们研究了通过喂养hfd在小鼠中造成严重肥胖的模型,并研究了生物素补充和通过益生元(果寡糖,FOS)调节肠道菌群的效果。事实上,人们已经知道FOS可以改善代谢状态,至少在一定程度上纠正由致肥性饮食引起的肠道菌群丰富度和组成。37-39在另一项单独使用FOS与HFD同时饲喂的实验中,我们发现与单独使用HFD相比,血浆生物素水平和生物素生产者丰度增加,考虑到总细菌丰度与否(在线补充图5C-E).尽管FOS对循环生物素水平有影响,但单独给药无法恢复附睾脂肪组织中生物素羧化酶的表达(在线补充图5A,讨论).
HFD导致肥胖(平均体重为37.9±5.1 g) 12周后,C57BL/6J成年雄性小鼠继续饲喂HFD,并在饮用水中添加FOS和/或在饮食中添加生物素,持续8周。在生物素补充组中,与未补充组相比,生物素循环水平增加了136.6倍(数据未显示)。在含有FOS和生物素的动物中,我们观察到体重增加显著受限,脂肪堆积减少(图6)、空腹血糖、空腹胰岛素下降和HOMA-IR改善(图6 b, C,在线补充图S7A).肠道菌群分析显示,分别补充FOS和生物素后,细菌多样性显著增加(FDR=5.05×10)−3与HFD组进行两两比较),FOS和生物素联合作用进一步增强(图6 d;罗斯福= 9.52×10−3).我们强调了生物素生产者相对丰度的显著增加和由生前补充介导的生物素转运体的显著减少(图6 e;罗斯福= 1.84×10−3和3.14×103,与HFD组)。单独补充生物素会导致携带生物素生物合成和运输基因的细菌群的特异性增加(图6 e;罗斯福= 2.82×10−2).同样,考虑到细菌的总丰度(数据未显示),也得到了类似的结果。生物素的补充本身可能促进特定生物素生产者谱系的增加以及微生物组的多样性。此外,考虑到细菌总丰度(数据未显示),其他B族维生素的生产者和转运者也同样受到生物素和FOS的补充的影响(在线补充图S7B).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
在小鼠实验中,添加生物素和FOS对既定肥胖的宿主代谢、生物素状态和微生物组组成的影响。(A)肥胖小鼠在82天(HFD 12周后和治疗前)到135天(FOS和/或生物素治疗8周后)之间的脂肪量增加(A: HFD+FOS (n=10) vs. HFD (n=5);B: HFD+FOS vs. HFD+生物素(n=9);C: HFD+生物素vs. HFD;D: HFD+FOS+生物素(n=5) vs. HFD;*P值<0.05,Kruskal-Wallis秩检验与Dunn 's多重比较检验)(B)使用FOS和/或生物素治疗6周后测定这些相同动物的空腹血糖(*P值<0.05,Kruskal Wallis秩检验与Dunn 's多重比较检验)。(C) FOS和/或生物素治疗6周后计算HOMA-IR指数(*P值<0.05,Kruskal Wallis秩检验与Dunn’s多重比较检验)。(D)长期肥胖小鼠不同组间Simpson多样性分布(**P值<0.01,FDR<0.05;配对Wilcoxon秩和检验)。(E)长期肥胖的不同组小鼠中生物素产生体(具有所有生物素生物合成基因但无生物素转运基因的细菌)、生物素转运体(无基因参与生物素生物合成的细菌)和生物素产生体+转运体(携带生物素生物合成和转运基因的细菌)的平均丰度(*P值和FDR配对Wilcoxon秩和检验<0.05)。 (F) mRNA expression of biotin carboxylases (ACCA, ACCB, MCC1, MCC2, PCCA, PCCB, PC) and biotin transporter SMVT in epididymal adipose tissue of mice with long term established obesity supplemented with FOS and/or biotin after 20 weeks of total follow-up (Kruskal-Wallis rank test, with Dunn’s multiple comparison; *P value and FDR<0.05, **P value and FDR<0.01, pairwise comparisons and P-trend were calculated using linear contrast tests). HFD, high-fat diet; SMVT, sodium multi vitamin transporter.
FOS和生物素的使用也诱导生物素羧化酶ACC1 (图6 f, p-trend = 1.50×10−2), ACC2 (p-trend = 2.39×10−3), MCC2 (p-trend = 2.13×10−2),表示(p-trend = 2.87×10−2)和生物素转运蛋白钠复合维生素转运蛋白(SMVT) (p-trend=6.38×10−3×10−2),以及PCCA呈上升趋势(p-trend=8.16×10−2)在附睾脂肪组织中。最后,当单独给药与肥胖的发展平行,独立于通过食物或皮下渗透微型泵给药的生物素给药模式时,观察到类似的结果。在线补充图S7C,D).
讨论
在细菌细胞丰度改变、低级别炎症和严重肥胖中出现的代谢改变等明显的生物失调的背景下,我们通过细菌生产者和营养不良菌的宏基因组量化评价了微生物生物素代谢的改变。严重的肥胖与微生物生物素的减少有关,这延伸到其他B族维生素。此外,宿主体内生物素利用率低,血液测量显示全身生物素水平低于理想水平。宿主生物素相对较低的另一个标志是脂肪组织中通过基因表达分析评估生物素依赖性羧化酶的表达降低。
由于在我们的人群中,通过膳食分析的生物素摄入量没有显示出主要的摄入差异,我们认为,转运蛋白的特别减少可能是由于生产者释放不足,他们自己也可能使用生物素或在当地细菌生态系统内。
人类宿主缺乏产生生物素的能力,因此生物素主要在空肠中从饮食中吸收,在较低程度上在远端肠中吸收。40我们在无菌和抗生素治疗小鼠上的结果表明,肠道菌群的缺乏对循环生物素水平有负面影响。
此外,在喂食HFD的小鼠中,收敛的实验结果强调了饮食质量在解释生物素代谢改变方面的重要性。致肥性饮食导致小鼠全身生物素减少,同时改变了微生物生物素代谢潜力。在人类群体中,细菌来源的生物素对循环生物素水平的贡献程度仍未测量。
生物素利用率低的起源也需要进一步的探索。相关研究揭示了微生物生物素代谢改变与肥胖相关代谢和全身炎症之间的相互作用。我们不能排除严重肥胖的代谢和炎症紊乱导致生物素细菌代谢改变。生物素通过肠上皮的运输也可能受到损害。这种转运需要宿主转运蛋白(SMVT (SLC5A6基因)。40在已发表的啮齿动物实验中,肠道炎症与生物素转运表达呈负相关。41tnf - α和干扰素- γ能够抑制体外培养的肠上皮细胞中SMVT的表达。41此外,LPS处理的结肠上皮细胞显示SMVT转运体的膜表达和结肠生物素摄取降低。42虽然我们无法在人类中探索这方面,但我们之前在严重肥胖的患者中发现,免疫细胞在空肠中积累,43生物素的主要吸收部位。在未发表的观察中,我们发现在严重肥胖患者中,空肠SMVT表达与CD4 +淋巴细胞和MAIT细胞的tnf - α表达呈负相关趋势,MAIT细胞是一种被肠道微生物活性激活的T细胞亚群。41因此,肠道炎症加剧,同时产生生物素的细菌数量减少,可能不利于宿主生物素的可用性。此外,考虑到生物素对脂肪组织稳态的重要性,不能排除在重度肥胖中,生物素的需求量在脂肪量大的过程中增加。由于肠道炎症导致的肠道生物素吸收减少,进一步放大了生物素对宿主的次优可利用性。有利于这一假设的是,BS同时诱导的代谢改善和炎症不仅导致了微生物生物素代谢的改善,而且还导致了全身生物素水平的升高。然而,由于经常吸收不良导致维生素补充不足,在BS和胃旁路手术中可以考虑更好地校正生物素状态。最后,在严重肥胖的情况下,宿主和肠道菌群可能会发生恶性循环,导致异常的代谢相互作用,来自不良菌群的分子信号可能导致宿主炎症和组织生物素缺乏的加剧。
因此,我们强调评估重度肥胖人群生物素缺乏的临床重要性。
我们意识到一些研究的局限性,这取决于生物素生产和微生物组消费潜力的宏基因组量化。宏基因组学数据本身并不能反映生物素生物合成和转运基因的真实生物活性,但根据参考基因和细菌种类的丰度,提供了微生物组实现这些生物功能的潜力的特征。元转录组学和可能的元蛋白质组学实验将有助于进一步量化微生物组对宿主生物素状态的功能贡献,并确定哪些细菌对生物素或其他维生素的循环水平很重要。
最后,基于我们结合小鼠和人类研究的结果,我们提出了一种肠道菌群失调的同时管理方法,即以肠道为中心的治疗(如益生元)和提高维生素的可用性,这看起来很有趣,可以防止肥胖过渡到更严重的代谢状态。因此,在安装肥胖的小鼠中,生物素和益生元一起给药可以限制体重增加和葡萄糖代谢恶化,同时改善微生物代谢。有趣的是,这种补充改善了生物素和其他B族维生素的细菌代谢。这为优化维生素B在肥胖中的地位提供了条件,并为未来在严重肥胖患者中应用生物素和益生元给药的临床研究铺平了道路。
方法
横断面队列描述(MetaCardis)
横断面欧洲MetaCardis研究队列由2214名参与者组成,这些参与者于2013年至2015年在法国巴黎Pitié-Salpêtrière医院的临床部门招募;在德国莱比锡的肥胖疾病综合研究和治疗中心(IFB)和丹麦哥本哈根的诺和诺德基金会基础代谢研究中心。排除标准包括年龄在75岁以上、腹部癌症/腹部放疗史、肠切除术史(阑尾切除术除外)、急性或慢性炎症或感染性疾病(包括丙型肝炎病毒VHC、V型肝炎病毒VHB和HIV)、器官移植史或接受免疫抑制治疗史、严重肾衰竭(肾病患者改变饮食(MDRD))肾小球滤过率<50 mL (min 1.73 m²)。−1),或者吸毒或酗酒。研究参与者在入组前3个月内不得使用任何抗生素。受试者提供书面知情同意,研究按照赫尔辛基宣言进行,并在临床试验中注册https://clinicaltrialsgov/show/NCT02059538.从粪便样本中获得了详细的生物临床信息以及肠道微生物宏基因组序列。在本研究中,我们根据BMI和葡萄糖代谢变化对1545名参与者进行了研究。
我们根据患者的BMI状况(非肥胖正常BMI (NOB BMI<25 kg/m²),超重/中度肥胖((MOB), BMI 25 - 34.9 kg/m²)),严重肥胖,(重度肥胖,SOB≥35 kg/m)2).此外,由于BMI在处理肥胖特异性共病方面存在一些限制,我们进一步根据代谢疾病的严重程度定义了三个不同的组,命名为“MH”(MH状态,由没有代谢综合征(MS)和/或T2D定义),“MUH”(MUH状态,由有MS定义,采用国际糖尿病联合会(IDF)的定义,但没有T2D),最后是“T2D”,以空腹血糖>6.9 mmol/L定义,和/或HbA1c≥6.5%和/或存在抗糖尿病药物。最终得到9个研究组。
代谢综合征的定义基于IDF 2005年代谢综合征全球共识定义(http://www.idf.org/metabolic-syndrome),例如,男性腰围>为94 cm,女性腰围>为80 cm,以及以下四个因素中的任意两个:(1)甘油三酯水平≥1.7 mmol/L或血脂异常的治疗(他汀类和/或贝特酸酯或依折麦布);(2)欧洲男性HDL胆固醇<1.03 mmol/L,欧洲女性< 1.29 mmol/L和/或脂质异常治疗;(3)血压异常:收缩压≥130 mm Hg和/或舒张压≥85 mm Hg或正在接受降压药治疗的患者;(4)空腹血糖异常≥5.6 mmol/L或T2D普遍。T2D状态的定义采用美国糖尿病协会的定义(https://care.diabetesjournals.org/content/41/Supplement_1/S13):空腹血糖>6.9 mmol/L和/或口服糖耐量试验2小时值>11 mmol/L和/或HbA1c≥6.5%和/或使用任何抗糖尿病治疗。
生化分析
对一夜禁食后采集的血液样本进行生化分析。采用酶法测定空腹血糖、总胆固醇、高密度脂蛋白胆固醇、甘油三酯和糖化血红蛋白,采用基于偶联酶系统的动力学测定法测定丙氨酸氨基转移酶、天冬氨酸氨基转移酶和γ-谷氨酰转移酶。根据弗里德瓦尔德方程计算法国和丹麦参与者的LDL胆固醇,测量德国参与者的LDL胆固醇。游离脂肪酸通过光度法测定(Diasys诊断系统)。化学发光分析法(Insulin Architect, Abbott)允许在空腹状态以及口服糖耐量试验中30和120分钟测量血清胰岛素和c肽,并使用人瘦素量化因子ELISA试剂盒(R&D Systems)测量血清瘦素。脂联素的测定采用ELISA夹心法(美国实验室产品公司(ALPCO)的高分子量和总脂联素ELISA试剂盒)。
使用image自动免疫分析系统(Beckman-Coulter)测定高敏感性CRP (hs-CRP)水平。使用人IL-6量化因子HS ELISA试剂盒(R&D Systems)测定血中HS -IL-6的浓度,使用人CD14量化因子ELISA试剂盒(R&D Systems)测定sCD14的浓度。使用Luminex测定法(ProcartaPlex Mix&Match Human 13-plex, eBioscience)测量以下细胞因子:干扰素- γ诱导蛋白10、C-X-C基序趋化因子配体5 (CXCL5)、CCL2、Eotaxine、IL-7、巨噬细胞迁移抑制因子、巨噬细胞炎症蛋白1β、基质细胞衍生因子1、血管内皮生长因子a。< 1个标准的值用1/2定量下限(LLoQ)代替,低于允许外推的下限的值用LLoQ/4代替。使用流式细胞术测量血细胞计数(白细胞、单核细胞、中性粒细胞和免疫细胞)。
在MetaCardis亚群的212个个体的血清样本中定量测定了生物素血清水平。样本是从健康状况极端的法国个体中特别选择的(NOB;n=107)和肥胖状态(SOB;N =105),也不包括有冠状动脉疾病的人。从8个研究组(NOB-MH, n=87, NOB-T2D, n=107, SOB-MH, n=86, SOB-T2D, n=101)共381个样本中,每组选取基因丰富度分布最极端的27个样本进行生物素血清定量。采用竞争性ELISA法测定血清样品中的生物素含量(MDbioscience,http://www.mdbiosciences.com/).用超高效液相色谱-串联质谱法测定尿排泄物中3-羟基异戊酸的含量。
人体测量学和临床数据的收集
在临床纳入访问期间,根据使用相同的量表和单位的标准化程序评估体重和身高。采用生物电阻抗法测定体脂质量和无脂质量。使用水银血压计测量收缩压和舒张压(每臂测量三次,右臂最后两次测量的平均值用于分析)。此外,还收集了一份详细的处方药物清单以及患者的病史。
生活方式和饮食数据的收集
身体活动的数据是通过近期身体活动问卷收集的。这份调查问卷评估了过去4周的身体活动实践,并对双标签水进行了验证。44饮食数据通过食物频率问卷(FFQ)收集,该问卷适应了每个招募国家的文化习惯。MetaCardis FFQ是基于经过验证的欧洲癌症前瞻性调查诺福克FFQ,内容基于几个相关的欧洲FFQ。分量大小和营养成分来源于全国食品消费调查和食品成分数据库。一项针对324名法国MetaCardis参与者的重复24小时饮食记录的验证研究表明,微营养素的平均相关系数为0.503,具有可接受的相对效度水平。45
生物素的摄入
虽然没有广泛的经过验证的生物素食物组成数据库,但我们根据两个独立来源的生物素含量值估计了生物素摄入量。第一个是斯塔格斯出版的精选食物等.30.第二项是根据最新的综合营养数据库fooddb (https://foodb.ca/)添加到丹麦国家食品数据库(DTU食品数据库:https://frida.fooddata.dk/)其生物素含量比1029种食物更完整。重要的是,我们发现基于FFQ的生物素摄入量估计是基于使用这两种方法得出的食物内容(已发表的值)30.与在线数据fooddb或DTU食品数据库)具有良好的相关性(平均相关系数为0.75)。因此,对于每个个体,我们决定将两种方法估计的膳食生物素摄入量取平均值,以调整我们的分析。这使得我们可以根据饮食中的生物素摄入量对个体进行排名,并调整我们对营养摄入的分析。
宏基因组测序
按照国际人类微生物组标准指南(SOP 07 V2 H)提取粪便总DNA,并使用离子质子技术(ThermoFisher Scientific)进行测序,平均每个样本的单端reads为23.3±400万(平均±SD) 150 bp。使用Alien Trimmer (V.0.2.4)清洗Reads,以去除弹性测序适配器,并修剪3 '侧的低质量核苷酸(质量和长度截止分别为20和45 bp)。清洗后的reads随后从人类和潜在的食物污染物DNA中进行过滤(使用人类基因组RCh37-p10、金牛和拟南芥,身份评分阈值为97%)。
粪便微生物负载
粪便样本的微生物量测定如前所述。17简单地说,0.2 g冷冻的(−80°C)等分物溶解在生理溶液中,总体积为100 mL (8.5 g/L NaCl;VWR国际(德国)。随后,将浆液稀释1000倍。样品用无菌注射器过滤器过滤(孔径为5µm;Sartorius Stedim生物技术,德国)。接下来,取1 mL微生物细胞悬液,用1µL SYBR Green I(1:100稀释DMSO;37°C荫蔽15分钟;10000浓缩液,赛默费雪科学公司,马萨诸塞州,美国)。使用C6 Accuri流式细胞仪(BD Biosciences, New Jersey, USA)进行流式细胞分析。46荧光事件监测使用FL1 533/30 nm和FL3 >670 nm光学探测器。此外,还收集了前方和侧面的散射光。使用BD Accuri CFlow (V.1.0.264.21)软件将FL1/FL3密度图上的微生物荧光事件与粪便样本背景进行门析和分离。在FL1通道上应用了阈值2000。门控荧光事件在正向/侧向密度图上进行评估,以排除其余背景事件。所有样品的仪器和门控设置保持一致(固定染色/门控策略)。46根据分析的等分物的确切重量,细胞计数被转换为每克粪便物质的微生物负荷。
宏基因组数据处理
使用人类微生物组的990万个基因集成参考目录进行了基因丰度分析。26使用METEOR软件中包含的Bowtie (v2.2.6)将筛选后的高质量reads以95%的身份阈值映射到990万个基因目录中。利用METEOR软件分两步生成基因丰度表。首先,唯一的映射reads(映射到目录中的单个基因的reads)被归因于它们对应的基因。其次,共享reads(映射到多个基因的比对得分相同的reads)根据其唯一映射计数的比例进行归类。使用MetaOMineR (V.1.2) R包对基因丰度表进行稀疏化、归一化和进一步分析。为了减少由于测序深度不同而造成的技术偏差,避免低丰度基因上的样本量伪产物,我们对读计数进行了细化。基因丰度表通过随机抽取1000万个不进行替换的映射reads,细化到每个样本1000万个reads。根据FPKM策略(通过基因大小和频率报告的总映射读取数进行归一化)对得到的精细化基因丰度表进行归一化,得到基因丰度谱表。
微生物基因丰富度(基因计数)是通过计算在给定样本中至少检测到一次的基因数量来计算的,使用10个独立稀疏实验中统计的基因的平均数量。
为了量化每个样本稀疏到10M reads后映射到IGC基因目录的功能模块宏基因组reads,按照MOCAT2框架中之前执行的IGC注释按功能类别分类(https://mocat.embl.de/).对每一类注释(如京都基因与基因组百科KEGG KOs,模块)的功能势在每个注释术语内进行相加。根据KEGG数据库(M00123, M00577, M00573)中生物素合成模块的平均丰度计算微生物组的生物素生产潜力。微生物组的生物素消耗潜力来源于KEGG模块M00581的丰度。
定量丰度分析
分类和功能宏基因组特征的相对宏基因组剖面(RMP)通过与样本微生物细胞密度成比例的索引因子(负载)的相对比例(定义为样本负载除以整个MetaCardis队列的中位数负载)相乘,转换为QMP。
Enterotyping
如Holmes所述,基于Dirichlet多项式混合物方法的肠分型(或群落分型)在R中执行,并使用包Dirichlet多项式等在n=2214个MetaCardis队列的一个子样本(n=2022)上(已有临床和宏基因组学数据)。
B族维生素生物合成和转运菌群的定量研究
为了对生物素和其他B族维生素的细菌代谢进行深入分析,我们根据Rodionov研究中定义的基因含量,根据生物素的生物合成和运输能力,对15个不同表型的细菌群的丰度进行了量化等.15简单地说,使用PATRIC命令行API从PATRIC数据库检索2126个参考基因组(https://www.patricbrc.org/).使用NGLESS将1545个样本中经过筛选的宏基因组与这2126个参考基因组进行比对。具有>95%身份和>45%对齐长度的Reads被保留,并使用NGLESS的dist1选项计算参考基因组丰度,其中首先从唯一的映射Reads计算基因组丰度,然后由由唯一映射Reads覆盖加权的多个映射Reads校正。利用MetaOMineR (V.1.2) R包对基因组丰度表进行稀疏化和RPKM归一化处理。原始基因组丰度表被细化到每个样本200万个reads,并根据FPKM策略(通过基因组大小和报告频率的基因组丰度进行归一化)对细化的基因组丰度表进行归一化。15个生物素物种群的丰度计算为单个细菌基因组丰度的总和。采用类似的方法计算生物素生产者(具有所有生物素生物合成基因的细菌,没有生物素运输基因)、生物素转运体(没有基因参与生物素生物合成的细菌)和生物素生产+转运体(含有生物素生物合成和运输基因的细菌)的总丰度。从这个丰度表(RMP)中,绝对丰度(QMP)通过将相对丰度乘以整个MetaCardis队列中样本微生物负荷与微生物负荷中值之间的比值来计算。
微生物多样性及群落结构分析
微生物基因丰富度(从IGC基因目录中具有非零丰度的基因数量)在每个样本将数据细化到1000万次读取后计算。不同疾病组间微生物基因丰富度的差异(不同肥胖状态的代谢健康组,按二甲双胍治疗分层的T2D个体;采用Kruskal-Wallis试验控制研究国家的潜在混杂因素(COIN R包的独立性测试功能(选项ytrafo=rank, teststat = ' quad ')进行评估。在每个比较中,所有研究组之间的两两差异用WRS检验对相同的潜在联合创始人进行调整(independence_test;options ytrafo=rank, teststat= ' scalar '),并使用Benjamini-Hochberg FDR进行多次测试校正。为了确定对微生物群变异具有非冗余解释能力的特征,首先对26个患者变量进行了属级群落排序(基于bry - curtis β -多样性矩阵的PCoA),包括疾病严重程度组、肥胖参数(BMI、脂肪量%、腰臀比)、代谢障碍的生物标志物(血糖、hdl -胆固醇、甘油三酯、HbA1c、尿酸)、代谢障碍的临床标志物(收缩期和舒张期血压、至少10%的纯素R包具有capscale功能的个体所摄入的药物。共识别出18个协变量(capscale p<0.05),随后用素R包的env2fit函数对这些协变量进行过滤,以识别具有非冗余解释力的协变量。
动物实验
GF小鼠粪便菌群转移实验
12-15周龄的雌性convi - r、GF瑞士韦氏鼠和C57BL/6 J鼠分别饲喂蒸压饲料(5021,LabDiet)和自由饮水。convi - r小鼠被安置在单独通风的笼子中(Green line Sealsafe plus, Tecniplast, Buguggiate, Italy),而GF小鼠被安置在柔性膜隔离器中;所有的老鼠都被关在一个笼子里,每个笼子最多5只老鼠,在严格的12小时光照周期下。通过厌氧和好氧培养粪便细菌,用通用引物27F(5’- gtttgatcctggctcap -3’)和1492R(5’-CGGCTACCTTGTTACGAC-3’)对细菌16S rRNA基因进行PCR检测GF状态。PCR反应在95°C预孵育5 min,然后在94°C 30 s, 52°C 45 s, 72°C 90 s,然后在72°C保存7 min,循环30个周期。
为了进行粪便菌群转移,将10-13周龄的雌性瑞士韦氏GF小鼠定植于两组有SOB和两组有NOB的小鼠的粪便中,为期4周,这两组小鼠的粪便中微生物菌群生物素的产生量高于SOB小鼠。小鼠被饲养在单独通风的笼子里(ISOcage N System, Tecniplast, Buguggiate, Italy),每个笼子最多5只老鼠。500 mg冰冻粪便悬浮在5 mL LYHBHI培养基(脑心灌注培养基,添加0.5%酵母提取物和0.1%纤维二糖、0.1%麦芽糖、0.05%半胱氨酸和0.25% hemin)中47含有20%甘油)。粪便泥浆在Coy室中制备,以1ml为容量,并在−80°C保存直到使用。小鼠随机分为4组,禁食4 h后灌胃2次,灌胃液为200 μ l;第二次灌胃于第一次给药后2天。来自每个供体的定植小鼠被自由喂养,要么是蒸压饲料(5021,LabDiet),要么是辐照的WD(40%千卡脂肪,TD.09683, Harlan Teklad)。
在禁食5小时后从腔静脉中采集血清样本。
利用QIIME2 (https://qiime2.org/).用QIIME2的DADA2插件对原始reads进行质量筛选、扩增子重建、嵌合体去除和扩增子序列变异的重建。利用QIIME2的Naïve贝叶斯分类器对GreenGenes 13_8数据库进行分类。原始ASV表被细化到每个样品30 000计数,以避免在检测稀疏曲线后样品间测序深度差异带来的偏差。用PICRUSt2对提纯ASV表进行了功能分析。48
动物
雄性C57BL/6 J小鼠来自法国Charles River实验室和Janvier实验室。成年雄性C57BL/6NTac来自Taconic Biosciences(丹麦)。小鼠被分成5组,保持12小时的光暗循环,并自由饮水。他们要么接受标准的chow饮食(A04-10,安全饮食,德国),要么接受HFD, D12492, 60%千卡脂肪,研究饮食,美国。这些实验是根据欧洲议会和理事会2010年9月22日关于保护用于科学目的的动物的指令2010/63/EU的建议进行的。实验得到了当地委员会的批准,并获得了法国“Ministère de la Recherche”的许可。
体重和组成
所有动物模型每周监测一次体重和食物摄入量。每周用核磁共振(Minispec LF50, Bruker, USA)评估全身成分,量化体脂、瘦组织和游离体液。在将小鼠插入仪器进行分析前称其体重(2分钟/只)。
抗生素管理
成年雄性C57BL/6NTac对照小鼠(n=16)每日灌胃用清水(n=8)或广谱抗生素溶液(100mg /kg万古霉素和200mg /kg氨苄西林、新霉素和甲硝唑)灌胃33用无菌水(n=8)稀释14天。
EGA手术
14只C57BL/6J动物行EGA。作为对照组,13只动物接受剖腹探查(假手术动物)。如前所述,执行EGA程序。35简而言之,接受手术的HFD或标准周氏饮食(A04-10,安全饮食,德国)的C57Bl6小鼠禁食6小时,并用2%异氟醚(Abbott, Rungis,法国)和空气/氧气麻醉。术前30分钟腹腔内给药(丁丙诺啡,0.03 mg/kg, Axience SAS,法国),手术结束时给药(酮洛芬,Merial, 1%,稀释1/100,150µL /只小鼠)。手术包括幽门括约肌结扎,然后进行肠胃吻合术,排除十二指肠和消化道近端空肠。假手术小鼠(简单剖腹手术)的麻醉时间与EGA小鼠相同。在两组动物中,剖腹修复分为两层。所有小鼠术后维持在标准化的治疗方案下,以监测疼痛、体重和水合,皮下注射生理盐水血清,必要时给予额外的镇痛。这些小鼠在手术后立即获得了水和食物。
生物素的补充
两组动物(雄性C57BL/6J, 8周龄)植入渗透泵(#2006,200µL, 0.15µL/hour, Alzet, USA),灌注生物素(Sigma Aldrich, USA)稀释浓度为75 mg/mL的载药溶液(水+5% DMSO;pH值调整到7.5)或单独的车辆。泵在异氟醚麻醉的小鼠皮下植入。
其他各组饲喂生物素补充的HFD定制饲粮,每公斤饲粮含有97.7 mg生物素(研究饲粮,美国)。
”丛书的治疗
在动物的饮用水中添加益生元(FOS, Orafti P95, Germany)。该化合物从Bénéo获得,并以10% w/w的剂量加入饮用水。以0.22 μ m过滤,每2天更换一次,避免变质。
空腹血糖和胰岛素测量
禁食6小时后称量体重,并从尾血中测定血糖。
用市售ELISA试剂盒(小鼠超敏胰岛素ELISA, ALPCO, USA)测定空腹6小时后采集的血样中的血浆胰岛素浓度,同时测定空腹血糖。qPCR在组织
使用RNeasy Mini Kit (QIAGEN,德国)从组织中提取总RNA。然后,使用随机引物和M-MLV (Promega, USA)反转录RNA。我们使用StepOne系统(Applied BiosystemsTM, USA)使用Fast SYBR Master Mix (Life Technologies, USA)进行qPCR。每个样本均归一化为管家基因18S和rplp0。利用该方程,用∆∆CT法计算基因表达的相对折叠变化2(-∆∆CT)。所使用的序列在表1在下面。
小鼠血浆/血清生物素定量
根据实验设计,在全身麻醉(3%异氟烷)下从眶后出血或在edta涂层管中禁食6小时后从尾静脉获得血液。然后将血液在13 000 g离心3分钟,收集血浆。在使用血清的实验中,在打开腹膜腔后,在麻醉(异氟烷)下通过腔静脉和门静脉插管采集血液。静置30分钟后,在4℃下1500 g离心10分钟,收集血清。根据IDK生物素ELISA试剂盒(K8141, immundiagnostics stickk)的说明,估计血浆/血清中的生物素水平。血浆/血清稀释至1:10和1:20。
粪便DNA提取
每周收集小鼠粪便,使用后立即在−80°C保存。然后,按照供应商的协议,使用PureLink微生物组DNA纯化试剂盒(Invitrogen)进行提取。用Qubit 4荧光计(Invitrogen)和NanoDrop (Thermofisher)评估DNA的数量和质量。
粪便DNA测序(纳米孔)
提取的粪便DNA由牛津纳米孔技术公司(ONT)的MinION测序。DNA文库使用连接测序试剂盒(ref SQK-LSK 109, ONT)制备,采用多路复用,最多可同时对12个样本进行测序(ref EXP-NBD104, ONT)。ZymoBIOMICS微生物群落DNA标准(#D6306, zyymo Research)被用作每次测序运行的内部控制。
粪便细菌丰度(qPCR)
将粪便DNA稀释到0.1 ng/µL,用StepOne系统(Applied BiosystemsTM, USA)和Fast SYBR Master Mix (Life Technologies, USA)对靶向16S rRNA基因的引物进行qPCR。
生物信息数据处理(纳米孔)
Nanopore fast5读取(平均每个样本343 206个读取)被基本调用并使用Guppy (V.2.1.3)进行多路复用。生物素和B族维生素细菌群的定量测定被用于将Nanopore fastq文件与上述来自Rodionov研究的2126个参考基因组进行比对等15使用minimap2校准器和map-ont选项优化的纳米孔测序数据。使用minimap2对齐的SAM文件产品作为NGLESS的输入,使用描述的dist1选项计算参考基因组丰度。利用MetaOMineR (V.1.2) R包对基因组丰度表进行稀疏化和RPKM归一化处理。原始基因组丰度表被细化到每个样本40000个映射读取,并根据FPKM策略(通过基因组大小和报告频率的基因组丰度进行归一化)对细化的基因组丰度表进行归一化。15个生物素物种群的丰度计算为单个细菌基因组丰度的总和。采用类似的方法计算生物素生产者(具有所有生物素生物合成基因的细菌,没有生物素运输基因)、生物素转运体(没有基因参与生物素生物合成的细菌)和生物素生产+转运体(含有生物素生物合成和运输基因的细菌)的总丰度。绝对宏基因组丰度是通过相对宏基因组丰度乘以16S rRNA基因的qPCR得到的细菌总丰度得到的。
来自独立BS队列的微生物组数据和生物素测量
我们量化了血清生物素,并从其他地方报道的涉及BS项目(Microbaria)的严重肥胖患者的粪便样本中进行了功能性宏基因组学。4简单地说,我们在巴黎Pitié-Salpêtrière医院营养科纳入了患有SOB的女性。根据国际BS指南,患者被分配为可调节胃束带或RYGB。在基线(T0)和术后1 (T1)、3 (T3)和12个月(T12)随访期间进行临床、人体测量学和生物学评价。每次来访都收集粪便。没有患者在BS发生前接受过3个月的抗生素治疗,既往无任何急性或慢性胃肠疾病史,干预前体重稳定。所有受试者都签署了知情的书面同意书,该方案在ClinicalTrial.gov上注册(Microbaria研究,NCT01454232).在基线时和BS后1年,对17例Microbaria队列患者的血清样本中生物素的血清水平进行了量化(共34个样本)。生物素测定采用竞争ELISA法,MDbioscience (http://www.mdbiosciences.com/).额外队列BS患者的生物素血清(BARICAN (CNIL No. 1222666;P050318 Les Comités de人身保护批准2006年11月24日,NCT01655017,NCT01454232)), (n=41例患者,每次T0:基线,T12:术后1年)采用IDK生物素ELISA试剂盒(K8141, immundiagnostic stik)检测。对未稀释和1:2稀释的血清进行测量。
生物素产生者和转运体的量化在24个来自Microbaria队列的个体中进行,在基线、1、3和12个月后的宏基因组数据。从ENA知识库中下载了这些个体的人类过滤SOLiD reads,并在上述Rodionov研究中提到的2126个参考基因组集合上进行了映射等15使用蝴蝶结允许最多3个不匹配和保持在每个层的最佳对齐。49SAM比对作为NGLESS的输入,使用上文所述的dist1选项计算参考基因组的丰度。利用MetaOMineR (V.1.2) R包对基因组丰度表进行稀疏化和RPKM归一化处理。原始基因组丰度表被细化到每个样本100万个reads,并根据FPKM策略(通过基因组大小和报告频率的基因组丰度进行归一化)对细化的基因组丰度表进行归一化。生物素产生体和转运体的丰度计算为每一组中单个细菌基因组丰度的总和,并将这些丰度乘以基因丰富度,以便根据微生物多样性和微生物细胞密度之间的正相关关系估算绝对宏基因组丰度(QMP)17在MetaCardis队列中观察(Spearman 's r=0.34, p=2.74×10)−44n = 1545人)。
脂肪组织基因表达与血液炎症标志物
对于43例接受BS的肥胖患者,皮下脂肪组织基因表达测量可通过微阵列和/或基于qPCR的方法获得(42例患者有微阵列数据,其中24例患者也有qPCR数据;25例患者有qPCR数据,其中1例患者没有可用的微阵列数据)。为了测量皮下脂肪组织样本的基因表达,从术前穿刺活检中提取RNA。在n=161名严重肥胖的受试者中,使用多重免疫测定法(基于多种规则的医学(RBM))测量血液炎症标志物。血液炎症标志物的缺失值(CCL2为n=8, IL-18为n=5, CEACAM1为n=4, CXCL9为n=3, ANGPT1为n=1, SPP1为n=1)使用LLoQ和beta定律进行计算。
微阵列实验
使用RNeasy总RNA迷你试剂盒(QIAGEN)单柱dna酶消化法从储存的组织中提取总RNA。使用安捷伦2100生物分析仪(Agilent Technologies)评估RNA质量和浓度。根据制造商说明使用Illumina RNA扩增试剂盒(Ambion)从250 ng总RNA中获得生物素标记的补充RNA。用Illumina Human HT-12 V.4.0 Expression beadchip (Illumina)进行杂交。杂化探针以cyanin-3-streptavidin (1 mg/mL;(Amersham Biosciences, GE Healthcare),并使用Illumina BeadArray阅读器进行扫描。原始数据通过genome estudio V.2011.1软件提取,使用默认设置,不进行任何额外的规范化处理。本研究重点选择的靶基因为生物素相关基因(SMVT、HLCS、BTD、ACACA、PCCB、PC、ACACB、PCCA、MCCC2)和炎症相关基因(A2M、ADIPOQ、ANGPT1、CAECAM1、CCL13、CCL19、CCL2、CCL23、CCL5、CCL8、CXCL10、CXCL12、CXCL9、DCN、FTH1、IGFBP3、igfb4、IGFBP5、igfb6、IL18、IL18BP、MMP2、MMP9、PECAM1、SERPINE1、SFTPD、SPP1、TIMP1、TNFRSF11B、TNFRSF1A、TNFRSF1B、TNFSF113B、VCAM1)。
qPCR实验
对于上述筛选的生物素相关基因,也采用qPCR检测BS前后皮下脂肪组织基因表达。对于每个用qPCR检测其表达的靶基因,通过减去基线时观察到的平均CT值(如BS前),将测量的CT值(如周期阈值)转换为ΔCTs。参考基因次黄嘌呤磷酸核糖转移酶(HRPT1)的CT值也以同样的方式转化为ΔCTs。然后用靶基因的ΔCTs减去HRPT1的ΔCTs,并对结果进行平方。为了在箱线图中更好地可视化,这些值还被转换为z分数。43,43
统计分析
所有统计分析采用R统计分析软件V.3.3.2进行。统计检验是两面性的。本文中所有报道的相关度量都是非参数斯皮尔曼相关。R2表示为%。
在适当的时候,使用Benjamini-Hochberg方法或Dunn方法(适用于小鼠模型)对所有p值进行多次测试校正。只有p值或FDR <0.05为显著性。关于模型的调整,在适当的时候,我们对招募国家和年龄进行了调整,因为它们被确定为肠道丰富度的混杂因素。对所有模型进行了关于混杂协变量的敏感性分析,并没有改变结果。
通过线性回归模型评估微生物组生物素生物合成和生产变量(来自IGC基因丰度和参考基因组)的对数转换绝对丰度与肥胖状态之间的关系,并考虑到该变量对微生物组组成的影响。5采用emmeans V.1.4.6 R包(Russell Lenth(2020))两两比较肥胖组间他定素调整线性回归的估计边际均值。emmeans:估计边际均值,又称最小二乘均值。V.1.4.6 R包。https://CRAN.R-project.org/package=emmeans),用Tukey法调整p值。
数据可用性声明
数据可在一个公共的、开放访问的存储库中获得。根据合理的要求提供数据。metacardishttps://www.ebi.ac.uk/ena/browser/view/PRJEB41311,https://www.ebi.ac.uk/ena/browser/view/PRJEB38742,https://www.ebi.ac.uk/ena/browser/view/PRJEB37249老鼠实验;16 s数据,https://www.ebi.ac.uk/ena/browser/view/PRJEB42967老鼠实验;纳米孔的数据,https://www.ebi.ac.uk/ena/browser/view/PRJEB42966,代码论文分析,https://git.ummisco.fr/ebelda/metatransitionbiotingut.git.
伦理语句
病人同意发表
伦理批准
该研究得到了法国伦理委员会、莱比锡大学医学院伦理委员会和丹麦首都大区伦理委员会的批准。研究设计(观察队列研究)符合所有相关伦理法规,与《赫尔辛基宣言》一致,并符合欧洲隐私立法。这些实验得到了瑞典哥德堡和法国巴黎索邦大学动物护理和使用伦理委员会的批准。所有动物实验均按照《实验动物护理及使用指南》的原则进行。
致谢
我们感谢受试者参与MetaCardis研究,特别感谢患者协会(Alliance du Coeur和CNAO)的输入和界面,以及Dominique Bonnefont-Rousselot博士(代谢生物化学科,Pitié-Salpêtrière医院)的血脂谱分析。我们还要感谢Manuela Krämer、Robert Jakubowicz和Anna Hallén在无菌小鼠和样品制备方面的工作。我们感谢Amélie Lacombe和PreclinICAN平台对小鼠实验的帮助。我们感谢来自心脏代谢和营养研究所临床调查平台的护士、技术人员、临床研究助理和数据管理人员进行的患者调查,感谢来自CRNH(法国营养研究中心)和来自Pitié-Salpêtrière医院的临床调查中心(CIC)的健康对照调查。广达医疗为临床研究提供监管监督,并为电子数据的处理和管理做出贡献。我们也感谢来自巴黎援助Publique-Hôpitaux (AP-HP)的支持,它是临床研究(MetaCardis和Microbaria研究)的促进者,SFN (Société Française de Nutrition),法国巴黎基金会和Artères, F-CRIN-FORCE网络的支持,以及通过ITMO提供的INSERM的支持。
参考文献
补充材料
脚注
推特@jeandbdt、@A_Myridakis @BackhedLab
EB、LV、VT、GF、SA和KEA的贡献相当。
JR和KC贡献相当。
调整通知这篇文章在Online First发布后进行了修改。作者姓名Edi Prifti已被更正。
合作者MetaCardis联盟合作者:罗希亚·阿利利,雷纳托·阿尔维斯,埃姆·阿斯特丽德·安德森·加利亚托维奇,奥利维尔·巴特勒米,让-菲利普·布德布里特,让-保罗·巴蒂斯,马格丽·伯兰,兰达·比塔尔,Hervé Blottière,弗雷德里克·博斯克特,拉希德·布布里特,奥利维尔·波隆,米克尔·加缪,多米尼克·卡苏托,塞西尔·西安古拉,让-菲利普·科莱,阿恩·迪特里希,莫拉德·杰巴尔,Angélique Doré,莱恩·恩格尔布莱希特森,利奥波德·费祖,塞巴斯蒂安·弗洛曼汀,菲利普·吉拉尔,玛丽安·格莱内,卡罗琳Grünemann,艾格尼丝·哈尔特曼,波莱特·哈特曼,杰拉德·海尔夫特,Malene Hornbak, Lesley Hoyles, Jean- sebastien Hulot, Richard Isnard, Sophie jaaqueminet, Niklas Rye Jørgensen, Hanna Julienne, Johanne Justesen, Judith Kammer, Nikolaj Karup, Mathieu Kerneis, Jean Khemis, Lars Køber, Ruby Kozlowski, Michael Kuhn, Aurelie Lampure, Véronique Lejard, Ivica Letunic, Florence Levenez, Lajos Marko, Lea Lucas- martini, Laura Martinez-Gili Robin Massey, Nicolas Maziers, Jonathan Medina-Stamminger, Lucas Moitinho-Silva, Gilles Montalescot, Sandrine Moutel,Ana Luisa Neves, Michael Olanipekun, Laetitia Pasero Le Pavin, Luis Pedro Coelho, Christine Poitou, Francoise Pousset, Laurence Pouzoulet, Andrea Rodriguez-Martinez, Sebastien Schmidt, Tatjana Schütz, Lucas Silva, Johanne Silvain, Mathilde Svendstrup, Timothy D Swartz, Thierry Vanduyvenboden, Camille Vatier, Eric O Verger, Stefanie Walther
贡献者EB、GF、JR和KC对项目进行了构思和设计。EB和GF提供了与患者表型相关的主要宏基因组分析,并调整了微生物细胞负荷。VT、LV、JD、TLR、CA和LM-H在小鼠体内进行研究,提供小鼠生物样品,并进行分析。EB为体内小鼠研究提供了宏基因组分析。KEA和SA确定膳食分析和模式以及与患者临床表型和炎症特征的关系,BH提供膳食FFQ数据的解释,SA和FM进行肠道炎症分析并解释患者炎症数据,VP提供人类组织中基因表达的分子分析,FA, MB, RC, JE-S, J-MO, JA-W, TN, KC和MS招募患者,RC, JA-W和TN有助于患者调查和数据管理。SH和PG提供了来自NutriNet-Santé研究的健康志愿者,EB, SF, ELC, GF, NP, EP, IL, JN, SV-S, BJ和PD开发数据库,分析管道,并进行宏基因组学和功能分析。BQ和HR负责粪便样本的处理和所有患者粪便样本的测序,CR、SAn和FM负责所有炎症细胞和系统标志物的测量,SFe负责代谢变量的集中测量。JC和AM对患者进行代谢组学测量,NBS提供网络分析以协助解释数据,J-DZ、SH、M-ED、JR、J-MO、OBP、PB、MS和SDE对结果进行讨论。EB、LV、VT、GF、SA、TLR、KEA、FB、JR、KC为论文作者。所有作者都对稿件进行了评论和编辑。 KC is the guarantor who accepts full responsibility for the work and/or the conduct of the study, had access to all data, and controlled the decision to publish.
资金这项工作得到了根据拨款协议HEALTH-F4-2012-305312开展的欧盟第七个研究、技术开发和示范框架方案(METACARDIS)的支持。同时获得Leducq基金会(17CVD01)、JPI-Microdiet study(2017-01996_3)的资助。部分工作得到了来自德国forschunggemeinschaft (DFG): SFB 1052(项目B1)、法国研究基金会Médicale (FDT201904008276, FDT202106012793)和法国研究机构(ANR-CAPTOR, anr - deepintegrgromics)的资助。
相互竞争的利益KC是Danone Research、Ysopia和CONFO therapeutics的顾问,其工作与本研究无关。KC在MetaCardis项目中与达能研究公司签订了合作研究合同。FB是Implexion pharma AB的股东之一。MB接受了阿斯利康、勃林格-英格翰、礼来、诺和诺德、诺华和赛诺菲的讲座和/或咨询费。
来源和同行评审不是委托;外部同行评议。
补充材料本内容由作者提供。它没有经过BMJ出版集团有限公司(BMJ)的审查,也可能没有经过同行评审。讨论的任何意见或建议仅仅是那些作者(s)和不被BMJ认可。BMJ放弃从放在内容上的任何依赖产生的所有责任和责任。如果内容包含任何翻译材料,BMJ不保证翻译的准确性和可靠性(包括但不限于当地法规、临床指南、术语、药品名称和药物剂量),并且不对翻译和改编或其他原因引起的任何错误和/或遗漏负责。
因为在这个版本中没有相关的实验,所以不得不删除这些句子
请求的权限
如果您希望重用这篇文章的任何部分或全部,请使用下面的链接,它将带您访问版权清除中心的RightsLink服务。您将能够快速获得价格和以多种不同方式重用内容的即时许可。