条文本

摘要
目标癌症的表观基因组改变与免疫微环境相互作用,决定肿瘤的进化和治疗反应。我们的目标是通过在胃癌中使用表观遗传交替启动子来研究肿瘤免疫微环境的调节,并将我们的发现扩展到其他胃肠道肿瘤。
设计交替启动子负载(APB)使用一种新的生物信息算法(高伦雅芙)从短读RNA测序和样品分类到APB中推断启动子活性高, APBint和APB低。单细胞RNA测序分析肿瘤内免疫微环境。一个人源化小鼠体内癌症模型被用来探索肿瘤动力学、替代启动子的使用和人类免疫系统之间的动态时间相互作用。采用免疫疗法治疗的多个胃肠道肿瘤队列评估了APB与治疗结果之间的相关性。
结果APB高胃癌肿瘤表现出t细胞溶解活性水平降低,并表现出免疫衰竭的特征。单细胞rna测序分析证实了APB中不同的免疫群体和较低的t细胞比例高肿瘤。使用具有活跃人类免疫系统的“人化小鼠”进行的体内功能研究显示,APB与肿瘤生长之间存在明显的时间关系高几乎没有人t细胞浸润的肿瘤。对免疫治疗的GI癌患者的分析证实了APB的耐药高肿瘤对免疫检查点抑制。APB高与APB相比,胃癌的无进展生存期明显较差低(中位数55天vs 121天,HR 0.40, 95% CI 0.18 ~ 0.93, p=0.032)。
结论这些发现证明了交替启动子的使用和肿瘤微环境之间的关联,导致免疫逃避和免疫治疗耐药性。
- 胃癌
- 肝细胞癌
- 免疫疗法
数据可用性声明
本手稿使用的数据包括先前发表的研究,基因组数据文件来自公共存储库:欧洲核苷酸档案:PRJEB25780和PRJEB34724癌症基因组图谱研究网络:dbGaP: phs000178.v10。p8本手稿中使用的数据包括先前发表的研究,基因组数据文件来自公共存储库:欧洲核苷酸档案:PRJEB25780和prjeb34724癌症基因组图谱研究网络:dbGaP: phs000178.v10。p8NanoString数据文件作为补充表(附件)提供。https://www.ebi.ac.uk/ena/data/view/PRJEB25780;https://www.ebi.ac.uk/ena/data/view/PRJEB34724https://www-ncbi-nlm-nih-gov.libproxy1.nus.edu.sg/projects/gap/cgi-bin/study.cgi?study_id=phs000178.v10.p8.
这是一篇开放获取的文章,按照创作共用署名非商业性(CC BY-NC 4.0)许可发布,该许可允许其他人以非商业性的方式发布、混编、改编、构建本作品,并以不同的条款授权他们的衍生作品,前提是原创作品被正确引用,给予适当的荣誉,任何更改都被注明,且使用是非商业性的。看到的:http://creativecommons.org/licenses/by-nc/4.0/.
来自Altmetric.com的统计
本研究的意义
关于这个问题,我们已经知道了什么?
免疫逃逸是肿瘤发生的关键因素。
癌症的表观遗传改变与免疫微环境相互作用,以控制肿瘤的发生和对治疗的反应。
胃癌的研究表明,表观遗传交替启动子、免疫编辑和免疫检查点抑制剂(ICI)耐药之间存在关联。
新的发现是什么?
具有较高表观遗传启动子改变的胃肿瘤显示t细胞溶解标记物水平降低,并表现出免疫衰竭的特征。
这些发现通过新的技术和平台如单细胞RNA测序和“人性化小鼠”进行了正交验证。
具有较高交替启动子负担的多种胃肠道肿瘤类型也与ICI治疗的较差生存率显著相关。
在可预见的未来,它会对临床实践产生怎样的影响?
替代启动子使用负担可能是适用于多种胃肠道肿瘤的免疫治疗的阴性预测生物标志物。
简介
在强大的宿主免疫监测下,肿瘤生长和转移是癌症的一个标志。免疫编辑是肿瘤细胞利用抗原呈递修饰、免疫检查点失调和免疫抵抗克隆选择等机制来逃避免疫识别的过程。1 - 3先前的研究表明,肿瘤可能通过吸收DNA甲基化、组蛋白乙酰化和染色质修饰等表观遗传机制来增强免疫编辑,以调节t细胞重编程、新抗原产生和免疫基因表达。4 - 7值得注意的是,由于表观遗传变化是潜在的可逆的,它们代表了抗癌治疗靶点的潜在节点。8
我们之前已经描述了胃癌(或胃腺癌(STAD))中使用替代启动子的免疫编辑新机制。9日10启动子是独联体-转录起始位点上游的调控元件,超过一半的人类基因有多个启动子。11启动子的活性受表观遗传学调控,使用备用启动子可以产生不同的5 '非翻译区和第一外显子,增强mRNA和蛋白质异构体的多样性。12通过表观基因组分析,我们证明了很大一部分STADs可以在高表达基因上使用交替启动子,产生缺失免疫原性n端肽的5 '截断蛋白异构体。交替启动子使用水平高的STADs被发现具有免疫安静表型,并可能对免疫检查点抑制剂(ICI)治疗产生耐药性。9日10
在这项研究中,我们试图研究胃癌中交替启动子的使用和肿瘤免疫微环境之间的关系,包括使用一种新的动物模型对肿瘤生长的时间影响。我们通过挖掘公共数据集和使用多重免疫治疗队列,将我们在胃癌中的发现扩展到其他胃肠道(GI)肿瘤。
方法
交替启动子负载(APB)算法
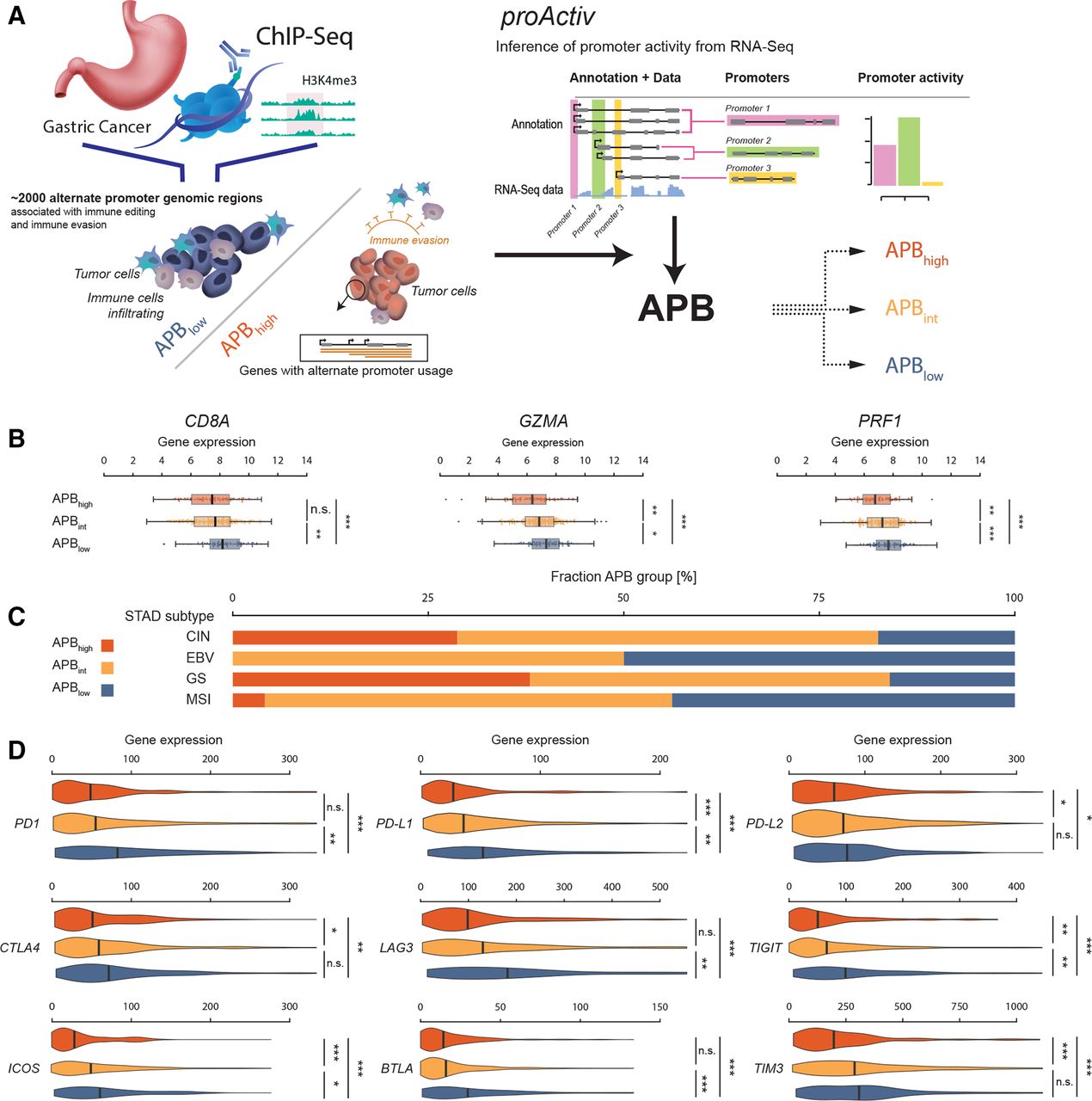
APB的计算公式包含高伦雅芙该算法通过绘制和量化基因组的第一个内含子连接,从短读RNA测序(RNA- seq)数据中估计启动子活性。高伦雅芙已被描述过,并可作为R包(在线补充方法).13将样本进行分组:APB的前四分位为APB高;最低的四分之一被归为APB低;其余样品为APBint(图1一个).在分组之前,对各批次/队列的APB水平进行归一化。

基于rna - seq的算法的发展,以衡量替代启动子的使用。(A)胃癌的表观遗传染色质免疫沉淀测序(ChIPSeq)研究确定了胃癌中与免疫编辑相关的特定表达获得(“获得启动子”)和表达丢失(“丢失启动子”)基因组区域。总共约有2000个备选启动子基因组区域被鉴定出来。高伦雅芙算法用于从短读批量RNA-Seq数据中推断启动子活性。高伦雅芙通过量化RNA-Seq转录本的第一个内含子连接推断启动子活性。从ChIPSeq数据中识别的基因组区域与高伦雅芙量化APB。每个队列内的样本根据APB进行分组高, APBint和APB低.(B) STAD中APB组与t细胞溶解活性标记物的关联(CD8A,GZMA而且PRF1).APB高组用红色表示,APBint黄色和全境通告低蓝色的。APB高与APB组相比,这3个基因的表达均较低int,这反过来又向APB显示了较低的表达低集团(Wilcoxon测试;* * * * * p < 0.001, p < 0.01, p < 0.05;n)。(C) APB组中STAD TCGA亚型的分布。STAD TCGA分子亚型:CIN、GS、EBV相关亚型和MSI。(D) STAD中APB组与9个免疫检查点的关联。与t细胞活性标志物相似,APB中这9个检查点的表达始终较低高(红色)与APB比较int(黄色)和APB低(蓝色)。(Wilcoxon测试;* * * * * p < 0.001, p < 0.01, p < 0.05;n)。APB,交替启动子负载;CIN,染色体不稳定;EBV, eb - barr病毒相关;GS,基因组稳定;MSI、微卫星不稳定性;n,不重要; STAD; stomach adenocarcinoma; TCGA, The Cancer Genome Atlas.
胃癌的单细胞rna序列
样本队列描述
在获得书面知情同意后,在新加坡国立大学医院接受手术切除的胃腺癌患者被纳入研究。
单细胞RNA测序(scRNA-Seq)文库制备
从肿瘤样本中分离出的单细胞在乳状液中生成凝胶珠,构建了丰富的5 '基因表达文库。经PCR扩增10×全长cDNA,从同一细胞中获得足够的材料构建多个文库。文库随后在Illumina Hiseq测序仪(在线补充方法).
人性化的小鼠模型
所有实验和程序均经A*STAR机构动物护理和使用委员会(IACUC, IACUC# 191440)根据农业食品和兽医局和新加坡实验动物研究国家咨询委员会的指导方针批准。
人类老鼠的一代
一至三天大的NOD-scid Il2rγ零(NSG)幼崽在1 Gy的亚致死辐射下移植1×105人类CD34+脐带血细胞(HLA-A24:02, Stemcell Technologies)通过肝内注射。人类免疫细胞重构超过10%的小鼠(根据人类CD45相对于人鼠CD45总和的比例计算)纳入研究。共选取5个HLA-A24:02亚型细胞系进行实验(2个APB高(sn1750和GSU),发出一份全境通告int(YCC21)及两份全境通告低(NCC 59和SNU16)细胞系)。
每个细胞系分别用5只人工小鼠和5只NSG小鼠注射肿瘤细胞,观察1个月。1个月后处死小鼠;例执行;肿瘤被收集以作进一步分析(在线补充方法).
Immunotherapy-treated临床军团
在一项多中心、产学研合作的研究中,收集了免疫疗法治疗过的GI癌症样本来评估APB水平。大部分样本来自不同组进行的各种ICI临床试验。贡献位点,肿瘤类型,ICI治疗和所进行的转录组分析类型(RNA-Seq vs NanoString)列于在线补充表S1.所有患者均在转移性、姑息性的情况下接受ICIs治疗。
泛癌症癌症基因组图谱(TCGA)分析
PanCanAtlas的基因表达数据和临床数据从Firebrowse下载。17Illumina HiSeq RNA-SeqV2 RSEM归一化基因值用于相关性CD8A,GZMA而且PRF1和其他免疫相关因素。数据库内的所有肿瘤类型均包括在内,但血液或免疫源性肿瘤除外(在线补充方法).
统计分析
定性数据的比例比较采用Fisher确切检验。参数量化资料比较采用双样本t检验。非参数定量资料比较采用Wilcoxon秩和检验。非参数双变量定量比较采用Spearman相关法。生存分析采用Kaplan-Meier曲线和log-rank检验。使用Cox比例风险回归模型评估hr和95% ci。所有分析均采用R V.4.0.5进行。
结果
基于rna - seq的替代启动子使用预测使泛癌症分析成为可能
H3K4me3 ChIP-Seq或Cap Analysis of Gene Expression tag测序是检测启动子活性的金标准技术。18然而,使用这些方法的数据通常只适用于小的肿瘤队列,而不适用于大型数据集或临床试验人群。为了克服这些限制,我们利用高伦雅芙,一种以前描述过的生物信息算法,从短读RNA-Seq数据推断启动子活性。13日19我们使用高伦雅芙通过先前的H3K4me3 ChIP-seq确定,使用一组预定义的备用启动子,相对于匹配的正常胃组织,对应于原发性STADs中获得或丢失的表观基因组区域,来估计肿瘤相关启动子的变化。9调节失调的交替启动子被识别高伦雅芙被量化,为每个肿瘤生成一个样本特异性的“APB”评分(APB) (图1一个).
首先,我们计算了TCGA中416个STAD样本的APB。STAD样本被归类为APB高: n=103 (25%), APBint: n=210(50%)和APB低: n = 103(25%)。APB高肿瘤的t细胞溶细胞标志物表达减少CD8A(CD8+肿瘤浸润淋巴细胞(TILs)),GZMA(granzyme)和PRF1(穿孔素1)(APB)高vs APB低, Wilcoxon检验,p<0.001;图1 b).20.作为阴性对照,与CD8A,GZMA而且PRF1当使用随机选择的大小相近的启动子子集计算APB时,未观察到表达(经验p<0.01)。这些结果与之前在STAD中的观察结果相似,支持使用高伦雅芙从RNA-Seq数据推断启动子活性。9日13
为了研究APB与四种TCGA STAD分子亚型(染色体不稳定亚型(CIN)、基因组稳定亚型(GS)、eb病毒相关亚型(EBV)和微卫星不稳定亚型(MSI))之间的关系,我们分析了376种TCGA亚型可分类的STAD。MSI和EBV亚型有少量/无APB高肿瘤(分别为4%和0%),而CIN和GS亚型的APB比例明显较高高肿瘤与APB比较低肿瘤(29% vs 17%, p<0.0001, 38% vs 16%, p=0.012) (图1 c).按Lauren的组织学亚型分类(即,弥漫性与肠道性)时,APB组间无显著差异(在线补充图S1A).APB组还与9个治疗相关性免疫检查点基因相关(PD-1,PD-L1,PD-L2,CTLA4,LAG3,TIM3,国际安全和发展理事会,BTLA而且TIGIT).所有免疫检查点基因在APB中表达均较低高肿瘤与APB比较低(9个基因均p≤0.01)(图1 d).排除MSI肿瘤后,APB之间的肿瘤突变负担(TMB)没有差异低和APB高肿瘤(3.6 vs 3.0 muts/Mb, p=0.12) (在线补充图S1B).
为了研究肿瘤基质成分的影响,我们将APB与从共识估计方法获得的肿瘤纯度相关联。21我们发现肿瘤含量与APB之间仅存在弱正相关(Pearson r=0.09, p=0.062) (在线补充图S1C).这些发现表明,肿瘤使用的替代启动子的估计不仅仅是由肿瘤样本中癌细胞的比例驱动的。接下来,我们将TCGA STAD样本的主要免疫细胞类型(来自CIBERSORT)的相对比例进行关联。22与之前的分析一致,APB低肿瘤有较高的T细胞丰度(CD4 (p=0.0068), CD8 (p=0.011)和T滤泡辅助细胞(p=0.033))和巨噬细胞M1丰度(p=0.039)。相比之下,APB高肥大细胞丰度较高(p=0.0079) (在线补充图S1D-H),提示先天免疫在APB中可能起作用高肿瘤。我们还将APB与ABSOLUTE计算的瘤内异质性评分相关。22APB高肿瘤似乎比APB有更高的瘤内异质性低肿瘤(p = 0.022) (在线补充图S1I).
scRNA-Seq证实了APB中不同的免疫群体高和APB低肿瘤
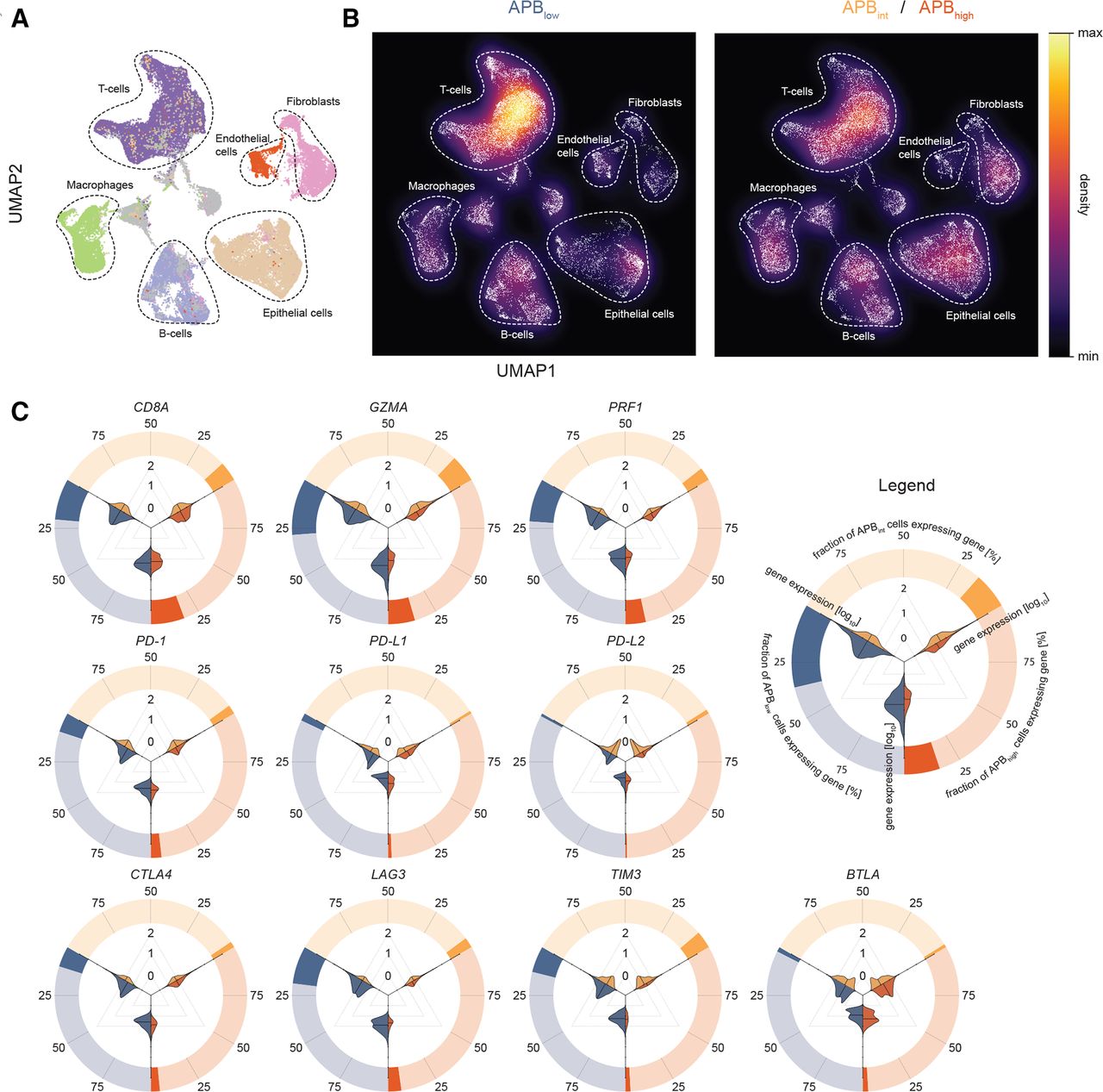
进一步调查是否全境通告高与APB相比,肿瘤具有明显或较少的肿瘤免疫细胞浸润低肿瘤,我们进行了scRNA-Seq。我们在11个手术切除的原发性STAD样本上生成配对的大体积RNA- seq和scRNA-Seq数据(55071个细胞)(由于RNA在异位特异性水平的测量低于当前scRNA-Seq技术的检测限度,只能从大体积RNA- seq中获得APB),将3个样本分类为APB低(19920细胞),5个为APBint(18 335个细胞)和3个APB高(16816个cell) (在线补充表S2).为了识别细胞群,基于基因表达谱进行无监督细胞聚类,允许维数减少到六种主要细胞类型:T细胞、上皮细胞、B细胞、巨噬细胞、内皮细胞和成纤维细胞(图2一个).

胃癌单细胞RNA-Seq及其与APB与肿瘤微环境的关系。(A) 11个样本55071个胃癌细胞的UMap(3个APB)高5 APBint3 APB低)以显示细胞的类型和簇。无监督分层聚类生成聚类,然后根据已知标记基因的表达进行映射和标记。主要的细胞类型包括上皮细胞(棕色)、T细胞(深紫色)、B细胞(浅紫色)、内皮细胞(红色)、成纤维细胞(粉色)和巨噬细胞(绿色)。(B)按APB分层的(A) UMap密度图低和APBint/ APB高突出显示APB中T细胞比例较高低APB中的肿瘤和上皮细胞int/ APB高肿瘤。(C)圆小提琴情节的表达CD8A,GZMA而且PRF1以及scRNA-Seq中APB组的免疫检查点。外圆表示表达该基因的APB组细胞的比例。例如,APB中~25%的细胞低肿瘤(蓝色)表达CD8A,而APB中不到25%的细胞高(红色)和APBint(黄色)肿瘤表达这种基因。里面的小提琴情节描述了APB组表达的程度。进行了三个比较:APB低(蓝色)和ABPint(黄色);APBint和APB高(红色)和APB低和APB高.从小提琴图中可以明显看出,在每个APB组中表达基因的细胞是相似的,然而,在APB中存在的这些细胞要少得多int和APB高肿瘤与APB比较低.这些结果表明,当CD8A-阳性细胞存在于APB中高和APBint肿瘤,低得多的GZMA而且PRF1都是由这些细胞表达的。APB,备用启动子使用负担;scRNA-Seq,单细胞RNA测序。
的表情CD8A,GZMA,PRF1在特定的细胞群中研究免疫检查点基因。CD8A,GZMA而且PRF1几乎只在T细胞中表达(约40%至55%的T细胞),证实了CD8A,GZMA而且PRF1作为t细胞溶解活性的合适替代物(在线补充图2A).同样,免疫检查点传统上被描述为在T细胞上表达,如PD-1,LAG3,TIGIT,TIM3,CTLA4而且国际安全和发展理事会T细胞中单细胞转录本表达显著升高(在线补充图S2A).我们比较了APB组之间的细胞比例(在线补充图S2B).总的来说,APB中t细胞比例较高低肿瘤与APB比较int/ APB高肿瘤(42% vs 30%, p<0.0001) (图2 b).相比之下,APB中的上皮细胞比例较低低肿瘤(13% vs 21%, p<0.0001)。值得注意的是,两组间b细胞组成无显著差异(14% vs 14%, p=0.82) (在线补充表S3).APB低肿瘤有明显的高表达CD8A,GZMA而且PRF1与APB相比高肿瘤(Wilcoxon p<0.0001),支持大量RNA-Seq分析。免疫检查点在APB中的表达也显著升高低肿瘤与APB比较高肿瘤(Wilcoxon p<0.0001) (图2 c而且在线补充图S2C),类似于大量RNA-Seq数据。总的来说,这些结果突出了不同的肿瘤免疫微环境和高、低交替启动子使用肿瘤之间的免疫检查点调控。
在人源化小鼠模型中交替启动子使用与免疫系统之间的相互作用
对原发肿瘤的分析通常只代表肿瘤的一个单一的时间快照,在手术切除或活检时获得。为了探索肿瘤动力学、替代启动子的使用和人类免疫系统之间的动态时间相互作用,我们使用了一个人源化小鼠体内癌症模型。NOD-scid Il2rγ零(NSG)免疫缺陷小鼠幼鼠移植HLA-A24:02人脐带血CD34+细胞和移植后人类免疫细胞重建的小鼠(称为“人性化小鼠”)被选择用于研究。5个HLA-A24:02亚型STAD细胞系(2个APB高,一个APBint和两个APB低细胞系)被选择(在线补充表S4).这些细胞株要么在市场上可以买到,要么从学术合作者那里获得。第23 - 25对于每一个STAD细胞系,5只人源化小鼠和5只NSG免疫缺陷小鼠在侧翼皮下注射肿瘤细胞。对产生的肿瘤进行生长速度、肿瘤大小、质量、体积和组织病理学分析(图3一).

APB的人源化小鼠模型研究。(A)拟人化小鼠模型实验研究方案。使用了5个细胞系(2个APB高,一个APBint和两个APB低细胞每一行)。每个细胞系分别用5只人源小鼠和5只免疫缺陷NSG小鼠的侧翼皮下注射肿瘤细胞,观察1个月。1个月后处死小鼠,进行尸检,取肿瘤进行分析。(B) APB组与NSG小鼠相比,人化小鼠肿瘤生长。青色线是人性化的老鼠;洋红色的线是NSG小鼠。APB高和APBint与NSG小鼠相比,人化小鼠的肿瘤生长更快,而在APB中低与NSG小鼠相比,人化小鼠的肿瘤生长较慢。(C)肿瘤生长的人化小鼠的肿瘤组织学。CD3和CD8免疫组化成像显示在SNU16 (APB低)(黑色箭头)和在SNU1750没有渗透(APB高).APB高肿瘤的CD3+和CD8+ t细胞浸润水平显著降低。黑条放大倍数50 μ m。(D) APB组人化小鼠肿瘤中免疫组化评分CD3和CD8的表达。用靶向CD3和CD8的抗体对FFPE组织进行免疫组化和H&E染色。显示任何强度的CD3或CD8明确染色的细胞百分比由对临床病理和生存信息一无所知的病理学家确定。表达CD3或CD8的TILs在肿瘤内被定义为癌细胞巢内的淋巴细胞和与肿瘤细胞直接接触的区域内被发现。TILs的定量是由各自TIL人群所占瘤内区域的百分比决定的。小提琴图突出了APB组(APB高(红色)和APB低(蓝色))。APB,备用启动子使用负担;直到,tumour-infiltrating淋巴细胞。
考虑到在免疫系统活跃的情况下研究肿瘤生长动力学的重要性,与人性化小鼠相比,NSG小鼠的肿瘤吸收和生长启动更快:97%的NSG和63%的人性化小鼠在第7天发生了肿瘤(Fisher的精确p=0.00056)。值得注意的是,APB高和APBint与APB相比,细胞系的肿瘤生长速度更快低人类小鼠的细胞系,与APB一致高肿瘤表现出免疫逃逸潜力,并在活跃的免疫系统中显示肿瘤发生。特别是在人化小鼠中,APB高90%的细胞系发生肿瘤(免疫缺陷小鼠为100%);APBint100%肿瘤发展(免疫缺陷患者100%);和APB低在第7天有20% (NSG中93%)的肿瘤发展(APB高/ APBintvs APB低, Fisher的精确p=0.039)。比较了NSG和人性化小鼠的每个细胞系的生长速率。对于APB高和APBint在细胞系中,人类小鼠的肿瘤似乎比NSG小鼠的肿瘤生长得更快,而APB则相反低NSG小鼠的肿瘤生长速度比人类小鼠快(图3 b).实验结束时,APB低在人类小鼠中生长的肿瘤比在NSG小鼠中生长的肿瘤小(中位数为375毫米)3.vs 512毫米3., p = 0.033)。相反,对于APBint和APB高细胞系,人源化小鼠肿瘤趋向于比NSG小鼠生长的肿瘤更大(中位数418毫米)3.vs 257毫米3., p = 0.073)。
1个月后处死小鼠,取肿瘤,镜检和免疫组化分析。APB低肿瘤显示明显的人t细胞浸润肿瘤,而APB高肿瘤几乎没有人t细胞浸润(图3 c).APB低肿瘤的CD3+(宽t细胞标记物)细胞浸润明显高于APB高肿瘤(15% vs 4%, p=0.0085)和CD8+(细胞毒性t细胞标记物)细胞浸润增加的趋势(10% vs 2%, p=0.088) (图3 d).接下来,我们对实验结束时从人化小鼠中获得的肿瘤进行了大体积RNA-Seq,研究标记特定细胞亚型的基因表达。APB低肿瘤表现出较高水平的适应性免疫细胞(CD8A而且PRF1(细胞毒性T细胞))和先天免疫细胞(PLD4(树突状细胞)和CD163(巨噬细胞))(在线补充图S3A).相比之下,APB高在人类小鼠中生长的肿瘤没有表现出这些特征,只有树突状细胞和巨噬细胞表现出较高水平(PLD4而且CD163,与先天免疫有关)(在线补充图S3B).除了免疫细胞类型外,我们还研究了人类小鼠肿瘤中免疫检查点的表达。APB低肿瘤表达了更高水平的LAG3, t细胞衰竭的标志TIM3(在线补充图S3C,D).这些来自人源化小鼠模型的发现表明APB高肿瘤很可能具有逃避免疫系统的能力,并具有免疫静音表型,这支持了原发肿瘤在体积和单细胞水平上的正交研究结果。
高替代启动子的使用与多种肿瘤类型对ICIs的耐药性相关
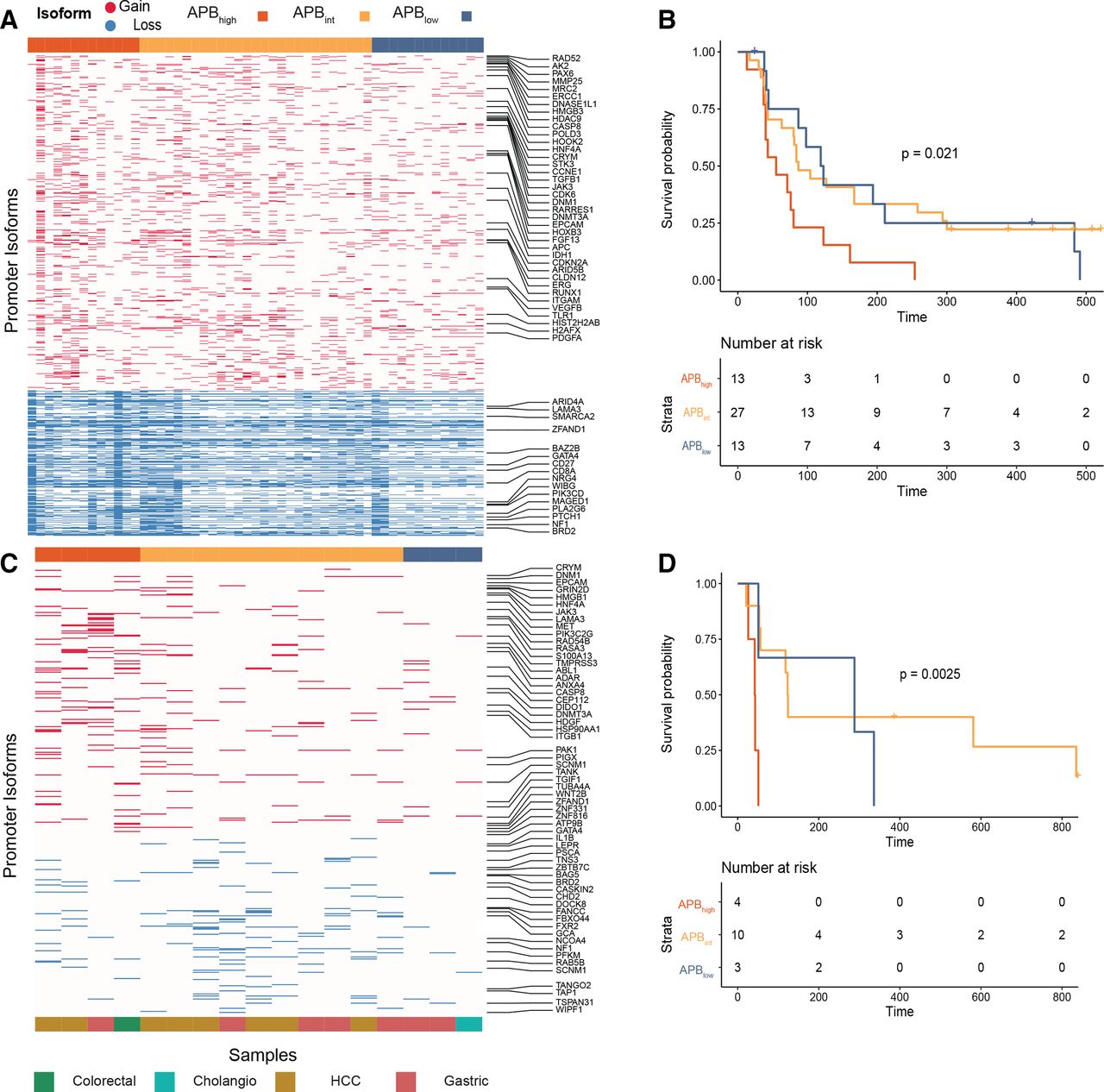
早期证据表明全境通告高STADs可能对ICI治疗有耐药性。10我们试图在这些发现的基础上进一步验证APB的假设高肿瘤对ICIs有抗性,因为它们的免疫安静表型。APB是根据53例使用pembrolizumab、nivolumab或atezolizumab(针对PD-1/PD-L1的单克隆抗体)治疗的胃癌样本的RNA-Seq数据计算的。中位年龄为57岁,75%为男性(在线补充表S5).队列中的所有患者均为转移性胃癌,在进入ICI临床试验前至少接受过一种系统性治疗。在53个样本中,13个肿瘤被归类为APB高;27人被列为全境通缉int;13例为通缉犯低(图4一).APB高与APB相比,肿瘤的无进展生存期(PFS)明显较差int(中位PFS 55 vs 87天,HR 0.39, 95% CI 0.19 ~ 0.80, p=0.01)和APB低肿瘤(中位PFS 55 vs 121天,HR 0.40, 95% CI 0.18 ~ 0.93, p=0.032) (图4 b).TCGA分型和PD-L1免疫组化数据分别为44例(83%)和40例(75%)(在线补充方法,表S5).EBV和MSI亚型肿瘤的PFS明显高于CIN或GS肿瘤(PFS未达到中位数vs 80天,HR 0.088, 95% CI 0.02 ~ 0.38, p=0.0011)。PD-L1联合阳性评分(CPS)≥10例患者的生存率也高于CPS <10例患者(中位PFS 254 vs 80天,HR 0.33, 95% CI 0.13 ~ 0.87, p=0.025)。这些数据与以前的发现一致。26日27日尤其值得临床关注的是在PD-L1 CPS <10和CIN/GS亚组中识别免疫治疗的预测性生物标志物。我们首先在PD-L1 CPS <10的亚组(n=31)中研究了APB,并发现了APB低和APBint肿瘤的PFS明显高于APB高(中位PFS 119 vs 84 vs 48天,HR 0.22, 95% CI 0.08 ~ 0.62, p=0.0042) (在线补充图S4A).当PD-L1 CPS <10的肿瘤仅局限于CIN/GS亚型时(n=28),结果相似(中位PFS 109 (APB低) vs 83 (APBint) vs 48天(全面通知高), HR 0.27, 95% CI 0.097 ~ 0.74, p=0.012) (在线补充图S4B).

APB阻力高肿瘤对免疫检查点抑制。(A)胃癌ci处理样品交替启动子使用热图。53个经ci治疗的胃癌样本(nivolumab, pembrolizumab和atezolizumab)用于本分析。获得替代启动子(在热图中标记为红色)和损失替代启动子(在热图中标记为蓝色)使用(列)每个样品(在行)。(B) APB组胃癌ICI PFS (n=53)。PFS与APB的Kaplan-Meier曲线高(红色)和APBint(黄色)和APB低(蓝色)。P值根据双侧对数秩检验。(C)使用NanoString对ici处理的非鳞状样品测量的交替启动子使用热图。总共,17个非鳞状icii处理样本的档案组织被用于NanoString分析,以计算APB。获得替代启动子(在热图中标记为红色)和损失替代启动子(在热图中标记为蓝色)使用(列)每个样品(在行)。(D) ci处理的非鳞状细胞样本的NanoString队列存活(n=17)。PFS与APB的Kaplan-Meier曲线高(红色)和APBint(黄色)和APB低(蓝色)。P值根据双侧对数秩检验。APB,备用启动子使用负担;ICI,免疫检查点抑制剂;PFS,无进展生存。
为了评估APB在预测对ICIs反应的特异性,我们下一步研究了APB在预测对其他类型全身治疗的敏感性方面的作用。我们分析了60例接受化疗(一线5-氟尿嘧啶和铂基化疗,在线补充表S1).三组间PFS无差异(中位PFS 158 (APB高) vs 129(全面通告int) vs 154天(全面通告低), p = 0.9) (在线补充图S4C).为了将此扩展到另一种全身治疗方案,我们随后测试了一组接受化疗和靶向治疗(紫杉醇和ramucirumab, n=47)联合治疗的胃癌独立队列患者。我们观察到两组间PFS缺乏相似的差异(中位PFS 104 (APB高) vs 128 (APBint) vs 126天(全面通告低), p = 0.8) (在线补充图S4D).这些发现表明,在所测试的治疗方法的局限性内,APB的预测性质可能是针对ICI治疗的。
对于绝大多数临床队列,包括来自临床试验的队列,只有存档的福尔马林固定石蜡包埋(FFPE)组织可用,这在RNA提取和使用RNA- seq表征方面提出了技术挑战。28NanoString平台已被验证可从FFPE组织中生成预测基因表达签名,并已用于临床实践。29为了探索更多的队列,我们设计了一个定制的NanoString面板,在NanoString(800个探针)的技术探针限制下推断FFPE组织中的APB。从用于计算RNA-Seq数据中的APB算法的4519个启动子中,我们从TCGA STAD分析中确定了排名最高的启动子,并为NanoString面板设计了备用启动子探针。纳米串探针被设计成主要与唯一的第一外显子结结合,允许识别和区分备用启动子转录本。这种方法在概念上类似于高伦雅芙识别增益和损失启动子转录本的算法13(在线补充方法).我们使用定制设计的NanoString面板测试了35个ci治疗的GI肿瘤的异质性队列。队列包括鳞癌(肛门和食道)(n=18)和非鳞癌(结肠、胃、胆管癌(CHOL)和肝细胞癌(HCC)) (n=17)组织学肿瘤。患者接受一系列ICIs治疗,包括nivolumab、avelumab和durvalumab(针对PD-1/PD-L1的单克隆抗体)。在非鳞状细胞队列中,APB高肿瘤的生存率低于APB低肿瘤(中位PFS 43 vs 288天,HR 0.10, 95% CI 0.01 ~ 0.78, p=0.028) (图4 c, D).类似的结果不能在鳞状癌队列中重复(中位PFS APB高241天vs全境通告int142天vs全境通告低216天,p=0.99)(见讨论部分)。为了进一步改进NanoString测量的APB (在线补充方法),我们对53例接受抗pd -1轴治疗的胃癌患者的档案FFPE组织进行了分析(在线补充表S1).用相似的公式推导出APB,识别出13个APB高, 20 APBint和20 APB低样本。PFS的APB高与APB相比明显降低int和APB低患者(48 vs 64 vs 175天,HR (APB高vs APB低) 0.25, 95% CI 0.11 ~ 0.57, p=0.001) (在线补充图S4E).
在展示了APB可潜在应用于其他GI肿瘤类型后,我们还研究了66个HCC样本的RNA-Seq数据,其中26个接受nivolumab治疗,40个接受pembrolizumab治疗。17个肿瘤被归为APB高;32例为APBint;17例为APB低.APB低肿瘤的PFS明显优于APBint/ APB高肿瘤(中位PFS 242 vs 90 vs 94天,HR 0.51, 95% CI 0.27 ~ 0.99, p=0.046) (在线补充图S4F).总的来说,这些发现表明我们对胃癌的初步结论可以扩展到高替代启动子使用的GI肿瘤,显示出对免疫检查点抑制的原发性耐药的更高概率,并提示使用APB作为免疫治疗的预测性生物标志物的作用。
高APB与多种肿瘤类型的免疫安静表型相关
最后,为了探索APB与多种肿瘤类型的免疫肿瘤微环境之间的关系,我们从TCGA PanCanAtlas RNA-Seq数据库中对26种肿瘤类型的10 165个样本的APB进行了量化。肿瘤被分类为APB高, APB低和APBint基于肿瘤类型特异性四分位数的分组(在线补充图S5A).与STADs相似,大多数肿瘤类型在APB组和CD8A,GZMA而且PRF1表达式。具体来说,在分析的26种肿瘤类型中,16种(62%)与所有3种标记物均有显著相关性(图5一个而且在线补充表S6).只有三种肿瘤类型与APB无相关性CD8A,GZMA或PRF1(肾上腺皮质癌(ACC), CHOL和葡萄膜黑色素瘤(UVM))。值得注意的是,后三个队列(ACC、CHOL和UVM)的样本量都相对较小(n<100,中位肿瘤类型数据集大小=414)。这些结果表明,APB与多种肿瘤类型的免疫微环境特征相关。

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
泛癌APB与免疫相关。(A)乳腺(BRCA)、结直肠、头颈部(HNSC)、肾脏、鳞状肺(LUSC)和黑色素瘤(SKCM)中APB组与t细胞溶解活性标记物的关联(CD8A,GZMA而且PRF1).的APB高组用红色表示,APBint黄色和全境通告低蓝色的。的APB高与APB组相比,这3个基因的表达均较低int,这反过来又向APB显示了较低的表达低集团(Wilcoxon测试;* * * * * p < 0.001, p < 0.01, p < 0.05;n)。(B) pananatlas中与APB相关的约20000个基因的火山图高和APB低6种肿瘤类型(BRCA,结直肠癌,HNSC,肾脏,LUSC和SKCM)。x轴是log2APB间基因表达的FC (RSEM)高和APB低。y轴是−log10调整后的p值结果(Bonferroni校正)。至少>1.5倍变化且调整p<0.01的基因为彩色,其余为灰色。过表达APB的免疫基因高为暗红色,而非免疫基因为淡红色。同样,APB低过表达的免疫基因为深蓝色,非免疫基因为淡蓝色。九个免疫检查点(PD1,PD-L1,PD-L2,LAG3,CTLA4,TIM3,国际安全和发展理事会,TIGIT而且BTLA贴上标签。免疫检查点在APB中过度表达是一种普遍趋势低肿瘤。APB,备用启动子使用负担;头颈部鳞状细胞;日志2FC、日志2褶皱变化;LUSC、肺鳞状;n,不重要;SKCM,黑色素瘤。
为了研究与高APB相关的其他免疫表型,APB之间的全基因组表达差异高和APB低肿瘤被发现。从总共约20,000个比较基因中,我们选择了import亚群——一个包含约4000个具有免疫相关功能和相互作用基因的数据库。30.大量的免疫基因在APB中表现出高表达低肿瘤相对于APB高几种肿瘤类型的肿瘤(阈值:倍变>1.5,调整后的p值<0.01)。例如,在乳腺癌中,导入基因组在APB中显著过表达低肿瘤(APB低: 731个基因vs APB高: 339, p≤0.0001)。所选免疫检查点基因的差异表达(PD-1,PD-L1,PD-L2,CTLA4,LAG3,TIM3,国际安全和发展理事会,BTLA而且TIGIT)在APB组之间也进行了研究。与STAD类似,APB中几乎所有免疫检查点都过表达低大多数肿瘤类型的肿瘤(图5 b).我们还利用了一项泛癌症TCGA研究,根据免疫特征对肿瘤进行分类。22APB低肿瘤具有更高的淋巴细胞浸润特征评分和干扰素-γ反应特征(p<0.0001) (在线补充图S5B、C).
7919个样本有MSI状态,其中176个肿瘤(2%)被发现为MSI高。APB组msi -高患病率为3.3%低, 2.5%的APBint和1.5% APB高(确切概率p = 0.0016)。因此,与STAD的发现相似,msi高的肿瘤似乎有被归类为APB的趋势低33%的msi高肿瘤为APB低相比之下,15%是APB高),尽管值得注意的是,在所有三个APB组中都发现了msi -高肿瘤。APB与TMB(中位APB)无相关性低1.8 vs APBint1.8 vs APB高1.8 muts/Mb, p=0.64) (在线补充图S5D).在MSI-H肿瘤中,APB与MSI-H之间TMB无差异高和APB低(p = 0.13)。总的来说,这些结果,基于基因表达分析在全球,免疫检查点和免疫信号水平,表明APB高肿瘤一贯与免疫“冷”肿瘤表型相关。
讨论
在这项研究中,我们试图探索替代启动子在癌症免疫和治疗反应中的作用。我们的研究结果扩展了先前的研究结果,将H3K4me3组蛋白谱应用于原发性STADs,以推断启动子活性,在STADs中反复丢失的肿瘤相关启动子异构体在高亲和性主要组织相容性复合体I类结合肽中显著富集。9我们提出,这可能反映了一种新的肿瘤免疫编辑机制,其中启动子的改变被用来逃避宿主免疫系统,从而促进新生肿瘤的发展。
替代启动子在癌症相关基因的使用,如切口已被描述在t细胞发育和致癌过程中发挥作用,而在ebv相关的自然杀伤t细胞淋巴瘤中,通过替代启动子的使用下调免疫原性病毒核抗原被认为是免疫逃逸的机制。31日32在我们的研究中,我们发现肿瘤在全基因组广泛使用替代启动子,这与免疫编辑和逃避的证据有关。值得注意的是,尽管使用了一组最初来源于STADs的启动子,但在多种肿瘤类型中观察到这些关联。替代启动子的使用可能与特定的起源组织关系较小,反映了对宿主免疫的保守泛癌反应。交替启动子使用、TMB和MSI之间的相关性表明,APB通过独立的机制途径与肿瘤免疫微环境相互作用,不同于DNA体细胞突变的获取(以TMB和MSI状态表示)。通过对APB与肿瘤含量和克隆异质性的相关分析,我们也证明了启动子的改变是通过多克隆机制驱动的。
为了从大量RNA-Seq数据中评估肿瘤微环境,我们使用了特定基因的表达,如CD8A,GZMA而且PRF1作为替代。使用单细胞RNA-Seq,允许识别和表征异质性肿瘤微环境中的不同细胞亚型,33我们通过确认这些基因的表达主要限于感兴趣的免疫细胞来验证这些替代标记的使用。我们的scRNA-Seq也描述了各种免疫检查点在STAD中按细胞类型的表达。这些发现,连同其他描述STAD免疫背景的发现,形成了开发细胞疗法和新的检查点抑制剂组合和策略的重要资源。34我们的scRNA-Seq数据集包括来自各种TCGA亚型的样本,包括CIN、GS和MSI。我们的研究的一个局限性是我们不能包括EBV胃癌,它与高水平的t细胞浸润和PD-L1表达有关。35EBV肿瘤往往更不活跃,在早期阶段被发现,并构成TCGA中最小的胃癌亚型(9%),在亚洲队列中发病率甚至更低(8%)。36我们研究中另一个有趣的发现是在APB中检测到更高水平的肥大细胞高STAD TCGA队列样本。肥大细胞是组织内固有免疫细胞,与适应性和固有免疫反应的激活和下调有关,37它们的活性可以由表观遗传调控因子调节。38肥大细胞通过OX40轴抑制调节性T细胞,从而改善效应T细胞的功能。39APB中肥大细胞较高高肿瘤可能反映了先天免疫的补偿性上调,因为T细胞水平较低(适应性免疫)。40
在本研究中,我们建立了一个体内模型来研究交替启动子使用与人体免疫系统之间的动态时间相互作用。在建立模型时,我们考虑了几个因素——首先,该模型需要功能免疫系统,因此禁止使用传统的免疫缺陷小鼠。其次,由于该模型必须允许对具有不同分子表型的肿瘤进行测试;传统的同基因小鼠模型,即肿瘤来自与宿主小鼠相同的遗传背景,被排除在外。人源化小鼠模型与传统的人类患者来源的异种移植模型(在免疫缺陷小鼠中生长)或体外细胞系/ t细胞共培养模型(用于测试肿瘤-免疫相互作用)之间的显著差距。41人源化小鼠模型已用于肝癌、肺癌、肉瘤和乳腺癌的研究。41 42我们的研究首次使用人源化小鼠分析性传播疾病,并采用与之前研究相对应的实验设计,比较人源化小鼠和免疫缺陷小鼠之间的肿瘤生长动力学。APB低与免疫缺陷小鼠和APB相比,人源化小鼠的肿瘤生长似乎更晚、更慢高肿瘤。在三阴性乳腺癌中也发现了类似的结果,与人性化小鼠相比,免疫缺陷小鼠的肿瘤生长得更快。42在一项肝癌研究中证实了在限制人类小鼠生长的肿瘤中细胞毒性T细胞的增加,使用派姆单抗治疗可进一步增加TILs。41值得注意的是,在我们的实验结束时,收集和分析肿瘤,以评估肿瘤免疫微环境,证明APB中缺乏TILs高肿瘤,这一发现与我们研究中的其他实验一致。本研究开发的人性化小鼠模型是测试治疗策略的理想平台,这些治疗策略的目标是肿瘤-免疫系统界面。然而,其局限性之一是,由于该平台此前未用于胃癌模型,因此使用免疫疗法和表观遗传药物对模型进行治疗将需要严格的优化和控制。小鼠模型实验的其他局限性包括皮下注射肿瘤。肿瘤的胃内移植已被描述为免疫缺陷患者衍生的类器官模型,并有可能被纳入未来的人源化小鼠胃癌实验。43 44
预后性生物标志物指导患者的预后或生存率,而预测性生物标志物提供了特定治疗的有效性的信息。45目前,最发达的预测ICI的生物标志物是免疫组化、MSI和TMB测定的PD-L1表达。46个47这些生物标记物是积极的预测生物标记物,可以识别可能对ICI产生反应的肿瘤。然而,围绕这些生物标志物的争议已经被提出,并且已经观察到生物标志物阴性人群中的ICI反应。48这些观察强调了ICIs的阴性预测生物标志物的补充作用,可以识别对治疗有耐药性的肿瘤,类似于拉预测抗egfr疗法耐药性的结直肠癌突变。49通过分析APB在化疗和靶向治疗队列中的预测价值,50 51我们证实,在所测试的治疗方法的局限性内,APB的预测性质可能是针对ICI治疗的。我们的研究结果可以通过将患者分为三组来指导选择进行免疫治疗的胃癌患者:第一,“可能的应答者”,包括MSI-H、EBV和PD-L1 CPS≥10个亚组。第二,“不可能应答者”,包括全境通告高患者,剩下的患者属于第三类“可能的反应者”。剩下的第三组患者倾向于从免疫治疗中表现出中等的益处,但有早期耐药性和中度PFS。
对免疫检查点抑制的敏感性是由各种因素驱动的,包括新抗原的形成,肿瘤突变负担和PD-L1的表达。47我们的胃肠道鳞状细胞癌队列包括食管癌和肛门癌,而我们的APB算法来自胃癌起源的腺癌样本。食管鳞状癌和腺癌组织学亚型的TCGA比较也发现了显著的基因组和表观遗传差异。52与食管腺癌相比,鳞状食管癌对免疫检查点抑制的反应比例也明显更高,这是由PD-L1的更高表达所驱动的。52 53派姆单抗对鳞状肛门癌也有类似的高应答率。54这些结果也反映在我们的队列中,在鳞状癌队列中,72%的疾病控制(稳定的疾病或部分反应)的患者是对ICI的最佳反应,而在腺癌队列中,这一比例为41%。因此,由于我们的APB算法似乎是一个阴性预测生物标志物的最佳功能,识别对ICI反应较差的鳞状癌,对ICI的高敏感性和较少的数量可能是纳米串小组未能区分鳞状GI癌队列中的应答者和非应答者的原因。对免疫治疗表现出原发性耐药的肿瘤亚组的识别与备选启动子相关,为生物标志物选择的联合治疗策略提供了有趣的可能性。靶向表观遗传途径的药物,如DNA甲基转移酶和组蛋白去乙酰化酶,已被证明可以通过靶向机制,如抗原处理和呈递、趋化因子和免疫检查点的表达和宿主免疫启动,消除免疫逃避。55目前正在进行几项试验,研究表观遗传因子与ICI结合的作用,旨在将免疫上的“冷”肿瘤转化为可能对免疫治疗更敏感的“热”肿瘤。55-57
总之,我们的研究描述了替代启动子的使用作为一种保守的泛癌症标志物,与免疫缺失的肿瘤微环境相关,而APB的量化可能作为免疫检查点抑制的一种新的阴性预测生物标志物。
数据可用性声明
本手稿使用的数据包括先前发表的研究,基因组数据文件来自公共存储库:欧洲核苷酸档案:PRJEB25780和PRJEB34724癌症基因组图谱研究网络:dbGaP: phs000178.v10。p8本手稿中使用的数据包括先前发表的研究,基因组数据文件来自公共存储库:欧洲核苷酸档案:PRJEB25780和prjeb34724癌症基因组图谱研究网络:dbGaP: phs000178.v10。p8NanoString数据文件作为补充表(附件)提供。https://www.ebi.ac.uk/ena/data/view/PRJEB25780;https://www.ebi.ac.uk/ena/data/view/PRJEB34724https://www-ncbi-nlm-nih-gov.libproxy1.nus.edu.sg/projects/gap/cgi-bin/study.cgi?study_id=phs000178.v10.p8.
伦理语句
病人同意发表
伦理批准
该研究获得了当地伦理委员会的批准(DSRB参考编号2005/00440)。
致谢
我们感谢患者和他们的家人参与我们的试验,并慷慨地同意使用组织来推进科学。
参考文献
补充材料
脚注
RS和PT是联合资深作者。
K-KH、VK、KR、DD和ZH贡献相等。
贡献者概念化:RS、AQ、JG、PT;数据整理:RS、KKH、DD、VK、KR、ZH、XO、ZFBAI、MX、AL-KT、DWMT、WZ、SPC、JQL、MDT、LM、EC、MF、MN、FP、JL、JY、CCYN、SYR、JS、KM、YN、WPY、QC、JG;形式分析:RS、KKH、AQ、VK、ADJ、MDS、YAG、AJS、JG、PT;获得资金:JG和PT;方法论:RS、VK、MDS、QC、DD、AQ、JG、PT;项目管理:RS和PT;资源及监督:QC, JG, PT;视象:RS、AQ、KKH、RP、AF;初稿撰写:RS;写作(审查和编辑):ZH, QC, JG, PT; approval of final version of manuscript: all authors.
资金RS得到新加坡国家医学研究理事会(NMRC)奖学金(NMRC/Fellowship/0059/2018和MOH-000627)的支持。FP得到了意大利sul Ricerca协会(AIRC IG 2019 Id.23624)的研究补助金支持。PT由杜克-新加坡国立大学医学院和生物医学研究委员会、科学、技术和研究机构支持。由国家研究基金会新加坡奖学金(NRF-NRFF2017-03)资助QC的人性化小鼠工作。这项工作也得到了国家医学研究委员会OF-LCG18May-0023、NR13NMR111OM和NMRC/STaR/0026/2015的资助。
相互竞争的利益本稿件的主题是作为技术披露提交给机构技术转让办公室,以潜在的知识产权保护。PT拥有HealthSeq的股票和其他所有权权益,从Kyowa Hakko Kirin和Thermo Fisher Scientific获得研究资金,并通过新加坡科学技术研究局获得专利/其他知识产权(所有在提交的工作之外)。RS获得了百时美施贵宝、礼来、罗氏、太和、阿斯利康、大昌华嘉和默沙东的酬金;曾与百时美施贵宝、卫材、默克、拜耳、泰禾、诺华、默沙东和阿斯利康开展咨询活动;接受帕克斯曼冷却器公司和默沙东公司的研究资助;获得阿斯利康、罗氏、卫材和太和制药(所有提交的作品之外)的旅行津贴。FP获得了来自安进、默克-雪兰诺、罗氏、礼来、赛诺菲、拜耳和Servier的演讲局/咨询活动酬金,并获得了BMS的研究资金(所有在提交的工作之外)。
来源和同行评审不是委托;外部同行评议。
补充材料本内容由作者提供。它没有经过BMJ出版集团有限公司(BMJ)的审查,也可能没有经过同行评审。讨论的任何意见或建议仅仅是那些作者(s)和不被BMJ认可。BMJ放弃从放在内容上的任何依赖产生的所有责任和责任。如果内容包含任何翻译材料,BMJ不保证翻译的准确性和可靠性(包括但不限于当地法规、临床指南、术语、药品名称和药物剂量),并且不对翻译和改编或其他原因引起的任何错误和/或遗漏负责。
请求的权限
如果您希望重用这篇文章的任何部分或全部,请使用下面的链接,它将带您访问版权清除中心的RightsLink服务。您将能够快速获得价格和以多种不同方式重用内容的即时许可。