
文摘gydF4y2Ba
增加膳食的摄入碳水化合物,在结肠发酵的微生物群报道降低体重,但机制尚不清楚。在这里,我们使用gydF4y2Ba在活的有机体内gydF4y2Ba11gydF4y2BaC-acetate醋酸和磁共振扫描显示结肠穿过血脑屏障,被大脑。腹腔内酯抑制食欲和下丘脑神经元激活模式。我们还表明,乙酸政府与乙酰辅酶a羧化酶的激活和表达谱的变化监管有利于抑制食欲的神经肽。此外,我们证明gydF4y2Ba13gydF4y2BaC高分辨率magic-angle-spinninggydF4y2Ba13gydF4y2BaC醋酸发酵的gydF4y2Ba13gydF4y2BaC-labelled碳水化合物在结肠内增加下丘脑gydF4y2Ba13gydF4y2Ba醋酸C高于基线水平。下丘脑gydF4y2Ba13gydF4y2Ba醋酸C区域增加了gydF4y2Ba13gydF4y2BaC glutamate-glutamine标签和GABA神经胶质的周期,与下丘脑gydF4y2Ba13gydF4y2BaC乳酸水平达到高于“剩余的大脑”。这些观察表明,乙酸在中央食欲调节有直接的作用。gydF4y2Ba
介绍gydF4y2Ba
肥胖已在世界范围内的通病,发病率20%以上在多数西方国家现在代表一个重大公共卫生负担gydF4y2Ba1gydF4y2Ba。这上升的肥胖是由于遗传易感性之间的不匹配的环境中生存的食物供应是有限的而当前肥环境,减少身体活动和能量摄入过剩gydF4y2Ba2gydF4y2Ba。现代食品加工导致的大规模生产廉价、高能量的食物,通常是高精制糖和脂肪,但低纤维gydF4y2Ba3gydF4y2Ba。据估计,旧石器时代饮食>每天100克的纤维,而当前西方摄入量每天只有10至20 ggydF4y2Ba4gydF4y2Ba。纤维在旧石器时代饮食消费,与当前的纤维摄入量,高度可发酵。越来越多的证据表明,链接发酵的膳食碳水化合物(包括纤维)结肠微生物群积极对代谢的影响gydF4y2Ba5gydF4y2Ba。观察由CanigydF4y2Baet al。gydF4y2Ba6gydF4y2Ba证明大鼠暴露于高脂肪饮食(HFD)增加可发酵碳水化合物(FC)组件4周较低膳食能量摄入、体重和肥胖,建议增加饮食FC导致改善食欲和体重管理。FCs被认为促进厌食的肠道激素释放肽YY (PYY组)和glucagon-like peptide-1 (GLP-1) (ref。gydF4y2Ba7gydF4y2Ba)。然而,FC-associated减肥在人类和啮齿动物模型未能持续显示增加这些食欲缺乏的激素gydF4y2Ba8gydF4y2Ba。gydF4y2Ba
我们最近所示小鼠膳食补充FC与减少能量摄入,体重,肥胖和下丘脑神经激活模式的变化,所有的独立的循环浓度的变化GLP-1 (ref。gydF4y2Ba9gydF4y2Ba)。为了更好地理解FC补充厌食的行动背后的机制,我们研究了最丰富的直接影响最终产品的结肠FC发酵,短链脂肪酸(SCFA)醋酸,控制食欲。gydF4y2Ba
我们现在这里的结肠发酵醋酸来源于FC行为直接抑制食欲通过下丘脑中部机制涉及transcellular神经递质变化周期。因此,开辟了新的研究方向为促进醋酸生产被结肠微生物群和治疗肥胖的预防和治疗策略。gydF4y2Ba
结果gydF4y2Ba
FCs对体重的影响gydF4y2Ba
我们给C57BL / 6小鼠HFD补充与FC菊糖(HF-I)或不佳可发酵的纤维素纤维(HF-C)。8周后,老鼠HF-I饮食获得大大减少体重比美联储HF-C (gydF4y2Ba图1一个gydF4y2Ba和消耗更少的食物gydF4y2Ba图1 bgydF4y2Ba)。SCFA结肠内容的分析前动物表现出显著增加总SCFA特别是最丰富的短链脂肪酸浓度乙酸(gydF4y2Ba图1 cgydF4y2Ba)。尽管其他人已经报道,FC补充导致增加食欲缺乏的激素GLP-1和肽yygydF4y2Ba6gydF4y2Ba我们组最近报道,FC-rich饮食增加manganese-enhanced MRI (MEMRI)信号在下丘脑,神经激活的一个代理,通过一个机制,似乎是独立于GLP-1变化(ref。gydF4y2Ba9gydF4y2Ba)。支持,厌食的肠道激素注入之前已经被证明能够抑制下丘脑MEMRI信号gydF4y2Ba10gydF4y2Ba的,从而表明增加信号后看到FC补充不是PYY组或GLP-1依赖。因此,我们假设有一个直接影响SCFAs食欲调节,即通过醋酸,它是最丰富的结肠产生的短链脂肪酸,这部分可以避免肝脏清除率达到微摩尔的末梢循环浓度。gydF4y2Ba

(gydF4y2Ba一个gydF4y2Ba)身体体重增加和(gydF4y2BabgydF4y2Ba)平均每周进食老鼠与一个HFD补充高度可发酵纤维菊粉(HF-I)或相对non-fermentable纤维纤维素(HF-C)。身体体重增加和平均食物摄入量显著降低HF-I集团* *gydF4y2BaPgydF4y2Ba< 0.01,* *gydF4y2BaPgydF4y2Ba基于双边< 0.05,未配对的学生gydF4y2BatgydF4y2Ba以及(gydF4y2BangydF4y2Ba= 12 /组)。(gydF4y2BacgydF4y2Ba)总acetate-only粪便短链脂肪酸浓度从HF-C和HF-I美联储获得老鼠* *gydF4y2BaPgydF4y2Ba基于双边< 0.01,未配对的学生gydF4y2BatgydF4y2Ba以及(gydF4y2BangydF4y2Ba每组随机选择从= 6gydF4y2BangydF4y2Ba= 24组)。(gydF4y2BadgydF4y2Ba)carbon-11 biodistribution (gydF4y2Ba11gydF4y2BaC)输液醋酸确定使用PET扫描。图片描绘了一个禁食鼠标后注入静脉输液。图像显示大脑中吸收、肝脏和心脏。(gydF4y2BaegydF4y2Ba- - - - - -gydF4y2BaggydF4y2Ba)大脑、肝脏和心脏的静脉输液和结肠注入gydF4y2Ba11gydF4y2BaC-acetate表示为一个百分比的初始剂量。相比没有显著差异,哎呀,Mann-WhitneygydF4y2BaUgydF4y2Ba以及,观察输液政府在美联储或禁食状态,但有一个缓慢增加的时候gydF4y2Ba11gydF4y2BaC-acetate colonically注入(gydF4y2BaPgydF4y2Ba< 0.001;gydF4y2BangydF4y2Ba每组)= 3 - 4。gydF4y2Ba
在大脑中醋酸的biodistribution和吸收gydF4y2Ba
SCFA传统上被认为是几乎完全由colonocytes和肝脏代谢gydF4y2Ba11gydF4y2Ba。然而,乙酸是末梢循环中,在人类的脑脊液gydF4y2Ba12gydF4y2Ba在大脑中,它为胶质细胞提供了一个重要的能量来源gydF4y2Ba13gydF4y2Ba。据我们所知,还没有以前出版调查醋酸作为一个潜在的代理量。因此我们试图调查醋酸的biodistribution外围地管理或管理它从FC colonically模仿内生生产。为此,我们使用gydF4y2Ba在活的有机体内gydF4y2Ba静脉注射(注射)和结肠的注入gydF4y2Ba11gydF4y2BaC-acetate pet - ct机和老鼠。pet - ct机代表禁食老鼠注射后的形象。gydF4y2Ba11gydF4y2BaC-acetate注入所示gydF4y2Ba图1 dgydF4y2Ba。肝脏和心脏演示了初始的最大比例gydF4y2Ba11gydF4y2BaC-acetate剂量摄取(gydF4y2Ba图1 f, ggydF4y2Ba),但我们的数据也显示,高达3%的醋酸最初由大脑静脉输液后注入示踪剂被美联储和禁食状态(gydF4y2Ba图1 egydF4y2Ba)。大脑吸收后结肠灌注是渐进的,但它~ 20分钟后达到同等水平。gydF4y2Ba
醋酸减少急性食物摄入量gydF4y2Ba
我们继续测试醋酸本身就是一个厌食的信号是否使用C57BL / 6小鼠急性外围政府。腹腔内(i.p)注射醋酸(500毫克公斤gydF4y2Ba−1gydF4y2Ba)醋酸生产增加血清(gydF4y2Ba图2一个gydF4y2Ba),类似于那些已经与游离脂肪酸抑制有关gydF4y2Ba14gydF4y2Ba相关,显著减少急性食物摄取1和2 h后注入(gydF4y2Ba图2 bgydF4y2Ba)。醋酸耐量试验证明没有影响醋酸的ip在500毫克公斤gydF4y2Ba−1gydF4y2Ba体重在血糖浓度(gydF4y2Ba图2 egydF4y2Ba)。急性行为观察注射后显示,这也是与厌恶无关的行为(gydF4y2Ba图2 dgydF4y2Ba)。此外,60分钟后注入,在循环血浆浓度没有明显差异PYY组GLP-1 (gydF4y2Ba图2 g, hgydF4y2Ba)。我们封装醋酸脂质体gydF4y2Ba15gydF4y2Ba。使用这种方法的外围管理liposome-encapsulated醋酸四健会期间,我们没有发现脂质体在下丘脑或食物摄入量的变化(gydF4y2Ba图2 fgydF4y2Ba)。因此,我们推测,醋酸的厌食的影响可能对中枢神经系统造成直接影响。我们测试这个假说的管理2.5μmol醋酸钠直接进入第三脑室intracerebroventricular (i.c.v)插管雄性Wistar鼠。依照厌食的醋酸外围地管理的影响,中央政府的醋酸第三脑室抑制食物摄取主要在1 - 2 h后注入,导致累积减少食物摄入量在2和4 h (gydF4y2Ba图2 cgydF4y2Ba),尽管这种影响是不一样的抑制小鼠醋酸ip管理后观察。喂养作为预期的短期方案考虑到快速肝代谢和醋酸的短半衰期。gydF4y2Ba

(gydF4y2Ba一个gydF4y2Ba醋酸)血清浓度的老鼠服用500毫克公斤gydF4y2Ba−1gydF4y2Baip醋酸或生理盐水(gydF4y2BangydF4y2Ba= 12)。(gydF4y2BabgydF4y2Ba)急性食物摄入数据显示食物摄入后小鼠急性醋酸ip管理(500毫克公斤gydF4y2Ba−1gydF4y2Ba)或生理盐水。在0 - 1和0 - 2 h食物摄入量显著降低;* * *gydF4y2BaPgydF4y2Ba< 0.001,* *gydF4y2BaPgydF4y2Ba< 0.01 (gydF4y2BangydF4y2Ba=每组21 - 22)。(gydF4y2BacgydF4y2Ba)2.5μmol醋酸钠的影响管理的第三脑室插管老鼠食物摄入量而氯化钠控制注入。交叉数据(gydF4y2BangydF4y2Ba= 15)相比,使用两面,成对的学生的gydF4y2BatgydF4y2Ba以及(gydF4y2BaPgydF4y2Ba= 0.05,gydF4y2BaPgydF4y2Ba= 0.08)。(gydF4y2BadgydF4y2Ba醋酸)小鼠的行为反应观察管理ip,盐水或氯化锂(积极的控制)。没有显著影响行为之间生理盐水或醋酸相比利用克鲁斯卡尔-沃利斯非参数方差分析(gydF4y2BangydF4y2Ba= 8)。(gydF4y2BaegydF4y2Ba)基线血糖浓度的变化(褶皱变化)后500毫克公斤gydF4y2Ba−1gydF4y2Ba醋酸钠或生理盐水注射gydF4y2Ba随意gydF4y2Ba老鼠(治疗效果gydF4y2BaPgydF4y2Ba= 0.6由双向方差分析;gydF4y2BangydF4y2Ba每组)= 9。(gydF4y2BafgydF4y2Ba乙酸)的影响ip liposome-encapsulated急性食物摄入量。在基于双边无显著差异,未配对的学生gydF4y2BatgydF4y2Ba以及(gydF4y2BangydF4y2Ba= 7)。(gydF4y2BaggydF4y2Ba,gydF4y2BahgydF4y2Ba)醋酸ip对等离子体浓度的影响肠肽PYY组GLP-1量。测量了60分钟后注入。没有显著影响肽是由两面,未配对,学生的gydF4y2BatgydF4y2Ba以及。gydF4y2Ba
醋酸引起的厌食的神经肽表达谱gydF4y2Ba
已经证实,乙酸是一个厌食的代理和确实是被大脑结肠或静脉输液管理后,我们下一个试图确认是否改变下丘脑神经元激活后,我们曾观察到FC的膳食补充剂gydF4y2Ba9gydF4y2Ba可以复制后急性醋酸。为了达到这个目标,我们再次使用了gydF4y2Ba在活的有机体内gydF4y2Ba功能成像技术MEMRIgydF4y2Ba9gydF4y2Ba,gydF4y2Ba10gydF4y2Ba,gydF4y2Ba16gydF4y2Ba。感兴趣的区域检查所示gydF4y2Ba图3gydF4y2Ba。小鼠的腹腔注射醋酸厌食的剂量的信号强度有显著增加的弓状核(ARC),表明增加神经元的激活,而没有观察到显著差异在下丘脑腹侧内侧(VMH),下丘脑室旁核(PVN) (gydF4y2Ba图3罪犯gydF4y2Ba)或脑干(gydF4y2Ba图3 egydF4y2Ba)。这些数据显示一个相同的模式来当我们看到美联储老鼠与FC菊粉HFD补充gydF4y2Ba9gydF4y2Ba。我们继续探讨醋酸特定ip后下丘脑神经肽的表达变化管理。有4倍增加肾上腺皮质的表达前体pro-opiomelanocortin (POMC)和一个强有力的抑制agouti-related肽(AgRP)表达式30分钟后醋酸管理局(gydF4y2Ba图3 fgydF4y2Ba)。在60分钟,抑制AgRP依然存在。这不同的表达模式的改变食欲缺乏的和orexigenic神经肽有利于减少食欲和体重,和之前报道了别人喂养啮齿动物FC-rich饮食gydF4y2Ba17gydF4y2Ba。gydF4y2Ba

(gydF4y2Ba一个gydF4y2Ba感兴趣的区域(roi)用于manganese-enhanced MRI (MEMRI)。先生的形象显示信号强度的ROI的位置在下丘脑(SI)测量确定。弧,弓状核;VMH、腹内侧下丘脑;ArP, postrema区域;nt,孤束核。白色的酒吧代表1毫米。(gydF4y2BabgydF4y2Ba- - - - - -gydF4y2BaegydF4y2Ba弧)下丘脑神经元激活,VMH、PVN和老鼠的nt ip管理醋酸或生理盐水后控制由MEMRI决定。箭头表示MnCl开始注射gydF4y2Ba2gydF4y2Ba输液。SI显著增加电弧的乙酸治疗小鼠相比saline-injected控制基于广义估计方程(GEE)和Mann-WhitneygydF4y2BaUgydF4y2Ba以及;* *gydF4y2BaPgydF4y2Ba< 0.01 (gydF4y2BangydF4y2Ba每组)= 4 - 5。(gydF4y2BafgydF4y2Ba)乙酸的影响下丘脑的表达POMC, NPY和AgRP由下丘脑qPCR决定。数据表示为褶皱的变化表达与saline-injected控制在30和60分钟后政府单向方差分析gydF4y2Ba事后gydF4y2BaDunnett校正;* *gydF4y2BaPgydF4y2Ba< 0.01,* * *gydF4y2BaPgydF4y2Ba< 0.001 (gydF4y2BangydF4y2Ba每组)= 5。(gydF4y2BaggydF4y2Ba,gydF4y2BahgydF4y2Ba扣带皮层部位和pAMPK下丘脑β-actin控制内容表达的关系,在双边基础上,未配对的学生gydF4y2BatgydF4y2Ba以及;*gydF4y2BaPgydF4y2Ba< 0.05 (gydF4y2BangydF4y2Ba= 5)。(gydF4y2Ba我gydF4y2Ba)免疫印迹老鼠的下丘脑pAMPK和扣带皮层部位水平30分钟后腹腔注射生理盐水或醋酸(完整的污点和注释中可用gydF4y2Ba补充图1gydF4y2Ba)。(gydF4y2BajgydF4y2Ba)提出了醋酸诱导的抑制机制喂养冲动。相对增加下丘脑柠檬酸循环活动增加ATP生产、减少安保水平,AMPK抑制乙酰辅酶a羧化酶(ACC),增加malonyl-CoA水平和刺激POMC mRNA表达,抑制食欲。gydF4y2Ba
醋酸降低下丘脑AMPK催化活性gydF4y2Ba
哺乳动物神经组织被认为迅速转化为乙酸,乙酰辅酶a,然后进入三羧酸循环gydF4y2Ba13gydF4y2Ba。越来越多的证据表明下丘脑活化蛋白激酶(AMPK)一个重要的角色在能源和营养传感gydF4y2Ba18gydF4y2Ba。172年苏氨酸磷酸化(T172)直接激活AMPK激酶的α亚基,进而磷酸化和乙酰辅酶a羧化酶灭活(ACC),导致malonyl-CoA水平下降。研究表明,增加下丘脑malonyl-CoA浓度与POMC的表达增加有关gydF4y2Ba19gydF4y2Ba和抑制神经肽Y (NPY)和AgRP随后啮齿动物食物摄入量的减少。醋酸因此,我们推测,政府可能调节食欲的变化我们也观察到,更具体地说POMC和AgRP表达的变化。我们测量的关键残基磷酸化AMPK (T172)和ACC (S79)在小鼠下丘脑ip后溶解产物醋酸注入的代理活动。我们发现水平显著减少pT172 AMPK和pS79 ACC (gydF4y2Ba图3胃肠道gydF4y2Ba醋酸),这表明急性政府灭活AMPK,从而导致ACC活动增加。ACC活动增加可以提升malonyl-CoA可以刺激POMC和可卡因的表情——amphetamine-regulated成绩单和降低NPY AgRP导致减少食物的摄入gydF4y2Ba19gydF4y2Ba。因为这意味着醋酸的厌食的影响的政府通过改变下丘脑ACC和AMPK活动和下游神经肽表达的变化。显示了这种关系的图解说明gydF4y2Ba图3 jgydF4y2Ba。gydF4y2Ba
13gydF4y2BaC醋酸优先在下丘脑积累gydF4y2Ba
鉴于上述改变神经元的激活和神经肽表达,我们试图确定醋酸的下丘脑的代谢利用gydF4y2Ba13gydF4y2BaC高分辨率magic-angle-spinning (HR-MAS)。gydF4y2Ba图4gydF4y2Ba总结了结果(2 -下丘脑i.p代谢gydF4y2Ba13gydF4y2BaC)醋酸或gydF4y2Ba13gydF4y2BaCgydF4y2Ba2gydF4y2Ba醋酸来自结肠发酵(U -gydF4y2Ba13gydF4y2BaC)菊粉由填喂法。gydF4y2Ba1gydF4y2BaH-decoupledgydF4y2Ba13gydF4y2BaC HR-MAS光谱脑活检提供三羧酸循环的操作信息(TCA)和transcellular glutamate-glutamine和γ-amino丁酸(GABA)周期gydF4y2Ba20.gydF4y2Ba,gydF4y2Ba21gydF4y2Ba。在这里,我们实现了gydF4y2Ba13gydF4y2BaC HR-MAS方法探讨代谢标记[2 -gydF4y2Ba13gydF4y2BaC)醋酸或gydF4y2Ba13gydF4y2BaCgydF4y2Ba2gydF4y2Ba乙酸在禁食老鼠的下丘脑切片的“剩余的大脑”获得了比较。为此,脑代谢被捕后立即与集中微波(0分钟)或15到30分钟之后,随着ip管理(2 -gydF4y2Ba13gydF4y2BaC)醋酸或180分钟后胃内的(U -gydF4y2Ba13gydF4y2BaC)菊粉,和下丘脑切片手术分离其余的大脑,调查gydF4y2Ba13gydF4y2BaC标签样本。gydF4y2Ba图4一gydF4y2Ba描述了gydF4y2Ba13gydF4y2BaC HR-MAS光谱从代表老鼠的下丘脑(2 -后15分钟gydF4y2Ba13gydF4y2Ba醋酸C)管理,揭示广泛gydF4y2Ba13gydF4y2BaC公司和显示最相关的共鸣。插图显示了代表gydF4y2Ba13gydF4y2BaC HR-MAS谱(28-38件分),从一只老鼠的下丘脑,获得180分钟后(U -gydF4y2Ba13gydF4y2BaC)菊粉填喂法管理。下丘脑中明显的存在解决了对比谷氨酸C4 (gydF4y2Ba1gydF4y2BaJgydF4y2Ba45gydF4y2Ba= 51 Hz)、谷氨酰胺C4 (gydF4y2Ba1gydF4y2BaJgydF4y2Ba45gydF4y2Ba= 49 Hz)和GABA C2 (gydF4y2Ba1gydF4y2BaJgydF4y2Ba12gydF4y2Ba= 51 Hz)证明gydF4y2Ba13gydF4y2BaCgydF4y2Ba2gydF4y2Ba醋酸(U -结肠发酵产生的单位gydF4y2Ba13gydF4y2BaC)菊粉已经纳入下丘脑的代谢。gydF4y2Ba图4 bgydF4y2Ba显示时间的gydF4y2Ba13gydF4y2BaC公司从这些基质在醋酸C2,谷氨酸和谷氨酰胺C4,乳酸GABA C2和C3碳在下丘脑切片(浅灰色),以及其余的大脑切片(深灰色)。的gydF4y2Ba13gydF4y2BaC HR-MAS共振的醋酸C2与时间达到更高强度显著增加比其余的大脑下丘脑,揭示非均匀分布的脑乙酸水平与下丘脑优惠积累的衬底。此外,gydF4y2Ba13gydF4y2BaC Glu C4的共振,Gln C4和GABA C2也随时间显著增加内部和extra-hypothalamic结构,支持大量的代谢积累的gydF4y2Ba13gydF4y2BaC醋酸脑三羧酸循环和神经胶质的glutamate-glutamine或GABA transcellular周期(gydF4y2Ba图4摄氏度gydF4y2Ba)。探讨醋酸的浓度是否来源于在结肠发酵足够高,以明确标签这些代谢物在下丘脑(触发厌食的反应描述),我们使用(U -gydF4y2Ba13gydF4y2Ba菊粉。有趣的是,gydF4y2Ba13gydF4y2BaC在这些代谢物从胃内的(U -积累gydF4y2Ba13gydF4y2BaC)菊粉通常高于i.p。(2 -gydF4y2Ba13gydF4y2BaC)醋酸,露出一个高效发酵菊粉的小鼠结肠水平直接影响下丘脑的代谢。gydF4y2Ba

(gydF4y2Ba一个gydF4y2Ba)代表gydF4y2Ba13gydF4y2BaC(125.03兆赫)HR-MAS光谱(4°C, 5 kHz)老鼠的下丘脑禁食过夜,15分钟后i.p。[2 -gydF4y2Ba13gydF4y2BaC)醋酸管理局(500毫克公斤gydF4y2Ba−1gydF4y2Ba)。插图:代表gydF4y2Ba13gydF4y2BaC HR-MAS光谱(28-38件分)从overnight-fasted老鼠的下丘脑,180分钟后(U -gydF4y2Ba13gydF4y2BaC]菊粉管理局(100毫克)填喂法。(gydF4y2BabgydF4y2Ba)增加gydF4y2Ba13gydF4y2BaC并入醋酸C2, GABA C2, Glu C4, Gln C4和虫胶C3碳(意味着+其中。)剩下的下丘脑和大脑切片,0后,15日,30分钟i.p。[2 -gydF4y2Ba13gydF4y2BaC)醋酸(n = 18)或180分钟胃内的[U -gydF4y2Ba13gydF4y2Ba菊粉政府*gydF4y2BaPgydF4y2Ba< 0.05,* *gydF4y2BaPgydF4y2Ba< 0.01,* * *gydF4y2BaPgydF4y2Ba< 0.001 (n = 4)。(gydF4y2BacgydF4y2Ba)总结的影响[2 -gydF4y2Ba13gydF4y2Ba醋酸C]或[U -gydF4y2Ba13gydF4y2Ba菊粉政府下丘脑的代谢。细胞外(2 -gydF4y2Ba13gydF4y2BaC)或gydF4y2Ba13gydF4y2BaCgydF4y2Ba2gydF4y2Ba醋酸来源于血浆和脑脊液运输主要星形胶质细胞,在三羧酸循环中氧化代谢,标签星形谷氨酰胺和谷氨酸C4 C4交换与相应的神经元通过glutamate-glutamine循环池,只在GABA ergic标签GABA神经元。(2 -gydF4y2Ba13gydF4y2BaC)或gydF4y2Ba13gydF4y2BaCgydF4y2Ba2gydF4y2Ba乙酸也氧化再生神经细胞周期中,原始的虫胶C3共振。glutamate-glutamine和GABA周期glutamatergic和GABA ergic神经传递的支持,两个基本突触事件触发增加CagydF4y2Ba2 +gydF4y2Ba吸收和竞争MngydF4y2Ba2 +gydF4y2Ba吸收(MEMRI)确定的欲望冲动。gydF4y2Ba
13gydF4y2BaCgydF4y2Ba2gydF4y2Ba从U -醋酸gydF4y2Ba13gydF4y2BaC菊粉快贴上Glu C4、Gln C4和下丘脑的GABA C2比其余的大脑。事实上,没有检测到gydF4y2Ba13gydF4y2BaC公司在GABA C2或Gln C4可以在剩下的大脑切片,发现即使gydF4y2Ba13gydF4y2BaC纳入共同Glu C4前体在这两个地区很相似。我们相信,这是优先整合的结果gydF4y2Ba13gydF4y2BaCgydF4y2Ba2gydF4y2Ba醋酸和下丘脑的代谢与其余的大脑相比,表明Gln C4和GABA C2标签non-hypothalamic结构在稍后的时间会发生。gydF4y2Ba
值得注意的是,gydF4y2Ba13gydF4y2BaC GABA和gydF4y2Ba13gydF4y2BaC乳酸保持在一个相似的水平直到30分钟gydF4y2Ba13gydF4y2BaC GABA和gydF4y2Ba13gydF4y2BaC乳酸在下丘脑高于其余的大脑切片,表明相对增加gaba ergic神经传递和氧化gydF4y2Ba13gydF4y2BaC乳酸生产丙酮酸的回收途径在小鼠下丘脑食欲刺激gydF4y2Ba22gydF4y2Ba。gydF4y2Ba
为了评估潜在的贡献在总代谢物含量变化gydF4y2Ba13gydF4y2BaC公司资料,我们收购了gydF4y2Ba1gydF4y2BaH HR-MAS相同的活检用于光谱gydF4y2Ba13gydF4y2BaC HR-MAS收购,共振检测到gydF4y2Ba1gydF4y2BaH HR-MAS揭示总代谢物含量。gydF4y2Ba图5一个gydF4y2Ba显示了一个代表gydF4y2Ba1gydF4y2BaH HR-MAS光谱从活检获得的下丘脑,180分钟后(U -gydF4y2Ba13gydF4y2BaC)菊粉管理局(黑色痕迹),与一些最相关的代谢物共振突出和相应的LCModel叠加(红色痕迹)。我们计算乙酸的代谢物比值,GABA,谷氨酸、谷氨酰胺和使用总乳酸gydF4y2BamyogydF4y2Ba肌醇或总肌酸(Cr + PCr)共振作为内部参考。我们的研究结果显示,显著提高gydF4y2Ba13gydF4y2BaC公司在下丘脑醋酸C2, GABA C2、谷氨酸和谷氨酰胺主要来源于C4和虫胶C3gydF4y2Ba13gydF4y2BaC-labelled基质管理而不是从相对总代谢物含量的变化(gydF4y2Ba图5 b-fgydF4y2Ba)。值得注意的是,类似的总谷氨酰胺和总GABA水平被发现在大脑下丘脑和剩余活检后180分钟(U -gydF4y2Ba13gydF4y2BaC)菊粉管理局(gydF4y2Ba图5 c, dgydF4y2Ba)。确认越高gydF4y2Ba13gydF4y2BaC公司发现在下丘脑Gln C4和GABA C2,并非来源于相对总代谢物水平的变化在这两个地区。当使用总肌酸也获得了类似的结论(Cr + PCr)共振总代谢物含量作为内部参考。这个标准化允许的,然而,对于下丘脑中的肌醇含量的比较,剩下的大脑。无显著差异,伊诺/ (Cr + PCr)这两个地区之间的比率被发现,15或30分钟后(2 -gydF4y2Ba13gydF4y2Ba醋酸C),或180分钟后(U -gydF4y2Ba13gydF4y2BaC)菊粉。这些结果表明,之前报道的差异未经处理的小鼠下丘脑和hipoccampus之间的肌醇含量gydF4y2Ba23gydF4y2Ba不明显的贡献目前比较后剩下的下丘脑和大脑切片(2 -gydF4y2Ba13gydF4y2Ba醋酸C)或(U -gydF4y2Ba13gydF4y2BaC)菊粉。gydF4y2Ba

(gydF4y2Ba一个gydF4y2Ba)gydF4y2Ba1gydF4y2BaHR-MAS光谱从下丘脑代表活检(黑色)和叠加LCModel配件(红色)。(gydF4y2BabgydF4y2Ba- - - - - -gydF4y2BafgydF4y2Ba)相对gydF4y2Ba1gydF4y2BaH HR-MAS比率Ac /伊诺,GABA /伊诺,Glu /伊诺,Gln /伊诺和虫胶/伊诺(2 -后在增加时间gydF4y2Ba13gydF4y2Ba醋酸C)或(U -gydF4y2Ba13gydF4y2BaC)菊粉。虫胶,乳酸;Ac,醋酸,Glu、谷氨酸;Gln谷氨酰胺;GABA,γ-amino丁酸;Cr + PCr,肌酸+磷酸肌酸;进气阀打开,gydF4y2BamyogydF4y2Ba肌醇;n。r,不是解决;CRLB, Cranmer-Rao下界< 20%。酒吧图表代表平均值和标准偏差。统计学意义是评估使用双尾未配对学生学习任务。“剩下的大脑”占脑extra-hypothalamic组织。*gydF4y2BaPgydF4y2Ba< 0.05,* *gydF4y2BaPgydF4y2Ba< 0.01。gydF4y2Ba
总结,我们得出结论说,在电弧中观察到神经元激活gydF4y2Ba图3 bgydF4y2Ba使用MEMRI是因为醋酸和glutamatergic或相对增加gaba ergic在下丘脑神经传递,会导致二价离子吸收,在这种情况下锰gydF4y2Ba2 +gydF4y2Ba,因此MEMRI增加信号强度,我们总结示意图所示(gydF4y2Ba图4摄氏度gydF4y2Ba)。gydF4y2Ba
讨论gydF4y2Ba
在这篇文章中,我们提供了一个新颖的见解背后的机制完善的观察动物美联储与FC HFD补充减少能量摄入和体重增加gydF4y2Ba24gydF4y2Ba,gydF4y2Ba25gydF4y2Ba。在这一领域的大部分工作调查机制这些观察此前专注于俱乐部的潜在影响促进厌食的肠道激素的释放PYY组GLP-1 (ref。gydF4y2Ba6gydF4y2Ba)。然而,这些激素浓度增加人类难以复制gydF4y2Ba26gydF4y2Ba。从我们组之前的研究,我们发现,减少食物摄入量和体重下降可能发生独立的小鼠肠道激素的变化gydF4y2Ba9gydF4y2Ba,gydF4y2Ba24gydF4y2Ba。我们还观察到增加MEMRI下丘脑电弧信号,没有相关的增加信号强度在脑干,相反的,观察后政府的厌食的肠道激素GLP-1和肽yygydF4y2Ba27gydF4y2Ba。我们已经证明降低体重FC补充研究中观察到的似乎是介导的,至少部分由SCFA醋酸。急性腹腔注射醋酸减少能量摄入/ 2 h不影响行为和发生独立的外围GLP-1和肽yy的浓度的变化。感兴趣的是封装的醋酸脂质体,专门为外围允许交付,废除了食欲缺乏的小鼠醋酸的影响,而醋酸i.c.v.管理在老鼠,尽管仍然厌食的,显示了一个更温和、更延迟抑制食物摄取。缺乏brain-stem-induced信号强度的变化后ip乙酸乙酸不表明迷走神经介导的效果,因此效力的差异可能是由于物种的变异。我们也证明食欲改变不太可能,因为变化的可用性葡萄糖下丘脑,醋酸,并不影响循环葡萄糖浓度。我们还表明,乙酸管理通过静脉输液和结肠路线被大脑和醋酸首次来自发酵的碳水化合物在结肠内被大量比其他大脑中的下丘脑组织。MEMRI信号强度模式后醋酸注入也直接反映了动物与FC美联储gydF4y2Ba9gydF4y2Ba。此外,我们表明,乙酸外围政府导致增加POMC和减少AgRP表达式在下丘脑,表明醋酸有直接影响的下丘脑控制食欲。类似的观察了动物与FC美联储gydF4y2Ba17gydF4y2Ba。gydF4y2Ba
变化的关键残基磷酸化AMPK和ACC在老鼠的下丘脑溶解产物醋酸外围政府也提供了证据的一个潜在的机制,通过神经肽表达可能发生的变化。鉴于醋酸被下丘脑,在优先星形胶质细胞代谢,因为更高的活动的monocarboxylate转运蛋白与神经元gydF4y2Ba28gydF4y2Ba,gydF4y2Ba29日gydF4y2Banon-neuronal机制也可能涉及观察增加电弧激活。使用gydF4y2Ba13gydF4y2BaC HR-MAS全部下丘脑组织,我们能够证明gydF4y2Ba13gydF4y2Ba醋酸C ip注射和来自结肠发酵合并到glutamate-glutamine transcellular周期,增加乳酸和GABA标签,从而支持下丘脑glutamatergic或GABA ergic神经传递。相同的模型提出了发生在乙酸代谢酒精的摄入gydF4y2Ba30.gydF4y2Ba。谷氨酰胺的操作周期包括必要的信号通过ionotropic metabotropic谷氨酸突触后受体,导致细胞内钙增加gydF4y2Ba2 +gydF4y2Ba积累gydF4y2Ba22gydF4y2Ba和伴随MEMRI效果gydF4y2Ba31日gydF4y2Ba(gydF4y2Ba图4摄氏度gydF4y2Ba)。它最近被证明在下丘脑瘦素调节谷氨酸转运体表达星形胶质细胞有一个关联的POMC神经元兴奋性增加gydF4y2Ba32gydF4y2Ba。类似的机制可以发生醋酸政府后,可能解释的增加MEMRI信号中观察到的动物补充与FC或独自醋酸。我们假设下丘脑的增加gydF4y2Ba13gydF4y2Ba醋酸度随着时间的推移,与大脑的休息,导致下丘脑增加gaba ergic神经传递从丙酮酸和乳酸氧化生产回收途径gydF4y2Ba22gydF4y2Ba。氧化代谢的增加会增加ATP生产、减少AMP: ATP比例,从而减少AMPK活性,从而减少ACC的抑制。这在malonyl-CoA产生增加,刺激主要POMC神经元和gaba ergic神经传递,从而减少的欲望冲动gydF4y2Ba33gydF4y2Ba。gydF4y2Ba
膳食纤维的摄入量超过每天100克被旧石器时代的人,在一些猎人聚集部落gydF4y2Ba4gydF4y2Ba建议人类结肠适应大量摄入可发酵材料,将导致醋酸high-peripheral循环gydF4y2Ba34gydF4y2Ba。纤维摄入量下降或者,更具体地说,减少碳水化合物的摄入,可以随时在结肠发酵这样的一个因素似乎是一个逆相关的肥胖患病率。虽然研究表明,包含高度FC饮食可能是有益的调节食欲和体重,在独立生存的个体,遵循这样的饮食方案往往是由于胃肠道副作用或不适口性低FC-rich食品。内的工作强调论文表明,乙酸SCFA可能至少部分调解一些obesity-protective FC-rich饮食直接影响中枢神经系统,从而表明醋酸可能是有用的作为一个潜在的治疗肥胖的方法。这个假设并不排除有其他非特异性效应。gydF4y2Ba
总之,这里我们提供新颖的见解通过FC可能调解机制抑制食欲。通过研究短链脂肪酸酯的作用,产品在结肠发酵的碳水化合物,我们的证据表明,乙酸源自结肠产生厌食的信号在下丘脑电弧通过支持glutamate-glutamine transcellular周期,导致乳酸和GABA产量的增加。这些数据表明先前未知的中央机制通过FC的发酵产品和膳食纤维可以帮助控制体重除此之外的能量稀释和肠道激素释放。此外,它开辟了重要的新的可能性为体重管理可发酵底物的供应,以冒号(因此醋酸生产)可以修改。gydF4y2Ba
方法gydF4y2Ba
动物和治疗gydF4y2Ba
动物实验进行伦敦帝国学院除了高分辨魔角旋转(浓度)实验进行研究院Investigaciones基于“阿尔贝托溶胶”(IIB)。伦敦帝国学院的实验是伦敦帝国理工学院批准,和所有动物程序进行依照英国动物科学程序法案(1986)。IIB实验学院伦理委员会批准Investigaciones基于“阿尔贝托溶胶”(和满足国家的指导方针(r 53/2013)和欧洲共同体(2010/62 /问题)保健和实验动物的管理。除非另有说明,所有实验进行雄性小鼠C57BL / 6(6 - 8周大,查尔斯河,马尔盖特,英国),single-housed温度控制(研讨会gydF4y2BaogydF4y2BaC)和光照条件(12 h光暗周期;灯在时间07:00)。gydF4y2Ba
HF-I的影响与HF-C体重和能量摄入gydF4y2Ba
老鼠被随机分配到两个不同的组(gydF4y2BangydF4y2Ba= 12):HFD与纤维素作为控制(HF-C) HFD + oligofructose-enriched菊粉(协同)(HF-I)。协同是一个包含两个长期和短链fructo-oligosaccharides fructan-based准备。协同与单独HFD混合的比例1:9。两个等热量的饮食,每个包含相同的总能量为4.6千卡ggydF4y2Ba−1gydF4y2Ba,41.8%的能源来自脂肪。饮食是由添加纤维素等热量的。营养成分的饮食所示gydF4y2Ba表1gydF4y2Ba。饮食喂养gydF4y2Ba随意gydF4y2Ba8周各自组的动物。身体重量和食物摄入量每周测量三次。gydF4y2Ba
醋酸浓度测定血清和粪便gydF4y2Ba
SCFAs气相色谱法测定小鼠的结肠内容和血清中,刚扑杀。方法是改编自理查森gydF4y2Baet al。gydF4y2Ba35gydF4y2Ba短暂,盲肠的内容重μg(20 - 220),结合550μl PBS。样本涡混合1分钟,离心机,享年3000岁gydF4y2BaggydF4y2Ba10分钟,上层的收集。500μl上层清液,25μl内部标准和2-ethylbutyric酸添加给最终的浓度5更易与lgydF4y2Ba−1gydF4y2Ba。酸提取250年除了μl浓盐酸和1毫升乙醚涡混合1分钟紧随其后。3000年样本离心10分钟gydF4y2BaggydF4y2Ba和醚层被转移到一个单独的小瓶。gydF4y2BaNgydF4y2Ba甲基-gydF4y2BaNgydF4y2Ba-t-butyldimethylsilyltrifluoroacetamide (MTBSTFA;σ)添加(100μl)加热20分钟的80°C。gydF4y2Ba
气相色谱法进行第二惠普5890系列仪器配备火焰离子化检测器,分流/不分流进样注射器和十米级,0.18毫米ID和0.20μm df Rtx-1 (Crossbond 100%二甲基聚硅氧烷,英国泰晤士Restek)毛细管柱。注入器和检测器温度是275gydF4y2BaogydF4y2BaC列温度从63°C编程3分钟到190gydF4y2BaogydF4y2Ba每分钟10°C。氦作为载气(压头135 kPa)和注射(1μl)在分割模式(50:1分裂)。峰地区记录和所有后续数据操作完成后使用ChemStation软件(美国安捷伦科技)。外部标准乙酸,丙酸,n-butyrate iso-butyrate, n-valerate浓度和己酸酯是准备在25日,12.5,6.25,1.25和0.625毫米和丁酸乙酯作为内部标准浓度的100毫米。报道值归一化根据原始样本的重量。gydF4y2Ba
血清SCFA测量gydF4y2Ba
100 - 500μl整除的血清是透过30-kDa微分区系统(Vivaspin RC VS02H22过滤器,缝匠肌Inc .米西索加,,加拿大),离心,享年14000岁gydF4y2BaggydF4y2Ba在4°C 90分钟。内部标准溶液(25μl)由ethylbutyrate 100毫米和100毫米甲酸是无蛋白的添加到225μl滤液上层清液中2毫升Hichrom瓶(安捷伦科技、南Queensferry、英国)。每个样本测量SCFA 1μl注入5890系列二世GC系统(惠普,克劳利,英国)装有NukolTM Capilllary列(30米×0.53毫米×1.0μm SUPELCOTM分析、英国)和火焰离子化检测器。氦,载气的流量是14毫升mingydF4y2Ba−1gydF4y2Ba。头部被设定为10与分裂组织注入压力。运行条件:初始温度60°C, 1分钟;+ 20°C mingydF4y2Ba−1gydF4y2Ba到145°C;+ 4°C mingydF4y2Ba−1gydF4y2Ba到200°C, 25分钟。山峰是综合使用安捷伦ChemStation软件(安捷伦科技,牛津大学,英国)和短链脂肪酸单点内部标准量化的内容的方法。峰身份和内部响应因素决定使用1毫米校准鸡尾酒包括乙酸、丙酸和丁酸。gydF4y2Ba
磁共振分析醋酸biodistributiongydF4y2Ba
小鼠禁食过夜。动物收到gydF4y2Ba11gydF4y2Ba初C-acetate(~ 20兆贝可)PET扫描通过尾静脉输液(gydF4y2BangydF4y2Ba= 3)或通过油管colonically放置1厘米到直肠(gydF4y2BangydF4y2Ba= 4)。扫描的时候,动物麻醉isofluorane-oxygen混合2 - 2%。动物后被放置在微型PET / CT扫描(PET / CT(西门子),没有对比进行了CT扫描,其次是PET扫描最后跟着CT扫描对比Ultravist 370(拜耳、AG)、德国)注入。gydF4y2Ba
在扫描期间,动物是维持在1 - 1% isofluorane-oxygen混合和体温37°C。CT的x射线源80千伏峰值和500μA曝光时间200毫秒和各向同性103μm的决议。扫描由三床位置重叠了20%。CT扫描是用来解剖数据和衰减校正的目的。宠物系统是由64年镏oxyothosilicate基于探测器单元安排在四个连续的戒指,水晶戒指的直径16.1厘米和12.7厘米的轴上。每个探测器块是由一系列20×20的镏oxyothosilicate晶体耦合通过导光管位置灵敏光电倍增管。每个晶体是10毫米长,横截面积为1.51×1.51毫米。晶体间距是1.59毫米的轴向和横向方向。Inveon研究工作版本3(西门子)是用于数据分析。图像矩阵大小12×128像素大小0.776毫米gydF4y2Ba2gydF4y2Ba和切片厚度0.796毫米被two-dimensional-filtered反向投影重建二维正弦图的使用一个坡道过滤奈奎斯特频率为0.5。动态框架是用于重建20×3 s框架,8×30年代框架,5×60帧和10×300年代框架。衰减校正也用于正确的图像。被注册到PET图像重建CT图像。宠物感兴趣的信号所需的地区获得扫描期间(1 h)。注射剂量比例(% ID / g)计算通过使用下列方程(1)和(2),在那里;gydF4y2BaλgydF4y2Ba= 0.693 /gydF4y2BaTgydF4y2Ba1/2gydF4y2Ba,gydF4y2BaTgydF4y2Ba1/2gydF4y2Ba放射性核素的半衰期,gydF4y2Ba我gydF4y2BatgydF4y2Ba活动在一个给定的时间在哪里gydF4y2Ba我gydF4y2Ba0gydF4y2Ba活动开始,t是时间的流逝,CgydF4y2Ba宠物gydF4y2Ba是图像的对比和ID是注射醋酸的活动。gydF4y2Ba
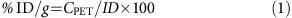

醋酸ip对能量摄入的影响gydF4y2Ba
小鼠禁食过夜之前接收500毫克公斤gydF4y2Ba−1gydF4y2Ba醋酸钠溶解在0.9%盐水(gydF4y2BangydF4y2Ba= 22)或车辆控制(0.9%生理盐水;gydF4y2BangydF4y2Ba乙酸= 21)pH值7.0,这个剂量是类似于之前的研究,用于抑制脂类分解并被证明是耐受性良好gydF4y2Ba14gydF4y2Ba。食物摄入量测量1,2,4 h后注入。gydF4y2Ba
醋酸i.c.v.对能量摄入的影响gydF4y2Ba
雄性Wistar鼠190 - 240 g(特定病原体免费;查尔斯河)被放置在一个立体框架(大卫·科夫仪器)在0.5 - -2.5%异氟烷麻醉。一个洞钻使用一个stereotactically电钻使用坐标计算从老鼠脑图谱如前所述gydF4y2Ba36gydF4y2Ba据Paxinos的坐标和沃森gydF4y2Ba37gydF4y2Ba(0.8毫米后的前囱中线和6.5毫米低于表面的头骨)。永久22码不锈钢套管(塑料公司,洛亚诺克,弗吉尼亚州,美国)投射到第三脑室移植。牙科水泥用于保存了套管的位置由三个不锈钢螺丝插入到头骨。皮肤是近似使用尼龙缝线。7天之后恢复期,动物每天处理,适应于注射过程。确保套管位置正确,老鼠收到i.c.v。150 ng血管紧张素ⅱ5-μl体积通过28-gauge不锈钢注射器放在和1毫米以下的套管。老鼠被观察到一个提示饮酒反应。5的总量μl意识注入超过1分钟,自由移动的老鼠。老鼠喂食研究(gydF4y2BangydF4y2Ba= 15)被禁食过夜,然后上午9点至10点小时注射醋酸钠(2.5μmol)或一个等价的渗透氯化钠控制(5μl),回到家中笼子。食物摄入量为1、2和4 h后注入。72 - h洗脱期后,老鼠收到醋酸或氯化钠以交叉的方式和食物摄入量测量如前所述。gydF4y2Ba
醋酸脂质体封装对食物摄取的影响gydF4y2Ba
纳米粒子的设计是基于我们之前的研究,聚乙二醇脂质体制定标签和可视化的细胞gydF4y2Ba38gydF4y2Ba,或者专门为优先吸收异种移植肿瘤gydF4y2Ba15gydF4y2Ba。脂质体是由薄膜水化法gydF4y2Ba39gydF4y2Ba。脂质体是准备用醋酸(1 M, pH值2.3)形成liposome-encapsulated醋酸(塔)纳米粒子或熟知的用于控制。颗粒大小是决定使用莫尔文Zetasizer(英国莫尔文仪器)。量化的乙酸纳米颗粒封装在塔决心使用gydF4y2Ba1gydF4y2BaH核磁共振(NMR)谱。LITA配方(200μl)是扫描与白蛋白的结合醋酸在自由溶液降低化合物的核磁共振信号gydF4y2Ba40gydF4y2Ba。liposome-free控制解决方案包含一个当量浓度的醋酸(4毫米)也扫描有和没有添加白蛋白(2 g)。利解决方案是扫描后的乳酸(5.2 mg乳酸钠)作为定量控制。光谱得到使用力量DRX 11.74 t NMR谱仪。光谱分析使用MestRe-C NMR光谱软件(西班牙圣地亚哥德孔波斯特拉)。gydF4y2Ba
评估短期biodistribution,包含一个额外的0.1%的脂质体配方rhodamine-lipid复杂(DOPE-Rhodamine: 1、2-dioleoyl-sn-glycero-3-phosphoethanolamine-N -(丽丝胺若丹明B磺酰)是用来确定biodistibution通过gydF4y2Ba体外gydF4y2Ba组织学分析。样本收集4 h ip后注入(gydF4y2BangydF4y2Ba= 4)。没有明显的积累脂质体在小鼠的大脑尽管存在于肝脏和心脏。gydF4y2Ba
醋酸的影响行为gydF4y2Ba
小鼠禁食过夜之前接收单个ip注入生理盐水,500毫克公斤gydF4y2Ba−1gydF4y2Ba醋酸或2.5氯化锂作为积极控制厌恶的行为gydF4y2Ba41gydF4y2Ba(gydF4y2BangydF4y2Ba每组= 8)。每个鼠标被观察到的行为模式每5分钟15秒的时间注入直到1 h后注入。行为分类之前使用修改后的版本的发布协议gydF4y2Ba42gydF4y2Ba。短暂,行为分为10个不同的类别:进食、饮酒、运动(包括饲养和攀登),床,穴居,打扮,不过,睡觉,头(动物体内异常的姿势:缩成一团,闭着眼睛半闭和pilo erect-indicative健康状况受损)和震动。gydF4y2Ba
醋酸耐量试验gydF4y2Ba
从血液葡萄糖水平测定来自老鼠尾巴使用一个精英Glucometer(拜耳公司)。实验进行gydF4y2Ba随意gydF4y2Ba美联储小鼠h 14时许为胰岛素耐量测试如前所述gydF4y2Ba18gydF4y2Ba而是胰岛素,动物被注射了ip与500毫克公斤gydF4y2Ba−1gydF4y2Ba醋酸钠或盐碱控制(gydF4y2BangydF4y2Ba= 9)。血糖值被确定在《纽约时报》表示。结果表示为褶皱初始注射前血糖浓度的变化。gydF4y2Ba
等离子体PYY组GLP-1激素分析gydF4y2Ba
所有样本化验的复制和在一个分析消除国米分析变异。等离子体PYY组GLP-1评估使用前面描述的建立内部等gydF4y2Ba43gydF4y2Ba,gydF4y2Ba44gydF4y2Ba。gydF4y2Ba
下丘脑定量聚合酶链反应gydF4y2Ba
这是使用比尤伊克描述的方法进行gydF4y2Baet al。gydF4y2Ba45gydF4y2Ba定量逆转录酶聚合酶链反应(RT-qPCR)被用来研究不同目标基因的表达。从整个下丘脑使用试剂盒提取的总RNA(英杰公司)根据制造商的协议。所有样本被DNaseI逆转录之前(表达载体)。第一链的互补是准备使用1μg RNA和SuperScriptII逆转录酶(表达载体)的随机六聚体和低聚糖(dT)引物(Promega、Charbonnieres-les-Bains、法国)。qPCRs是使用光周期计执行快速启动DNA主人SyBR绿色我工具包(罗氏,Meylan法国)在特定的引物的存在对选择放大的小片段(100 - 200个基点)。PCR产品检查特异性通过测量他们的熔化温度。样品(一式两份)量化相比之下的标准曲线得到稀释purified-specific互补。gydF4y2Ba
测量AGRP和POMC mRNA的表达gydF4y2Ba
RNA提取使用绝对从解剖下丘脑RNA从Stratagene microprep工具(La Jolla、钙、美国)。AgRP和POMC基因转录的下丘脑的电弧是决定使用实时rt - pcr和结果被表示为一个比还有本构基因的表达。还有TaqMan探针和引物的序列(加入基因库。M15933):正向引物5′-CCCACCGTGTTCTTCGACAT;反向引物5′-TGCAAACAGCTCGAAGCAGA-3′;和探针5′-CAAGGGCTCGCCATCAGCCG-3′。AGRP,他们是:正向引物5′-TTGGCAGAGGTGCTAGATCCA-3′;反向引物5′-AGGACTCGTGCAGCCTTACAC-3′;和探针5′-CGAGTCTCGTTCTCCGCGTCGC-3′。探针和引物POMC(分析识别。 Rn00595020_ml) were purchased from Applied Biosystems.
Manganese-enhanced核磁共振gydF4y2Ba
MEMRI执行使用9.4特斯拉瓦里安INOVA成像系统(美国瓦里安公司)如前所述gydF4y2Ba46gydF4y2Ba。快速旋转回声摘要多层序列使用以下参数:应用TR = 600毫秒,TE = 10 ms,矩阵大小= 256×192,FOV = 25×25毫米,平均收购46 = 1×0.4毫米厚片。一批66收购成立,46片整个注入了66倍。归一化比例增强信号强度计算弧,VMH, PVN,室旁核(PE)和原子核的束solitariusgydF4y2Ba27gydF4y2Ba,gydF4y2Ba43gydF4y2Ba。gydF4y2Ba
下丘脑腺苷酸激酶的测定gydF4y2Ba
动物被斩首,大脑被立即解剖和下丘脑和snap-frozen在液态氮。冷冻组织(~ 100毫克)均质在0.4毫升的冰冷的50 mm玫瑰,pH值7.4,50 mm氟化钠,5毫米焦磷酸钠,250毫米蔗糖,EDTA 1毫米,1毫米二硫苏糖醇,苯甲脒1毫米,1毫米胰蛋白酶抑制剂,0.1% (w / v) phenylmethylsulfonyl氟化物使用UltraTurax均质器(3×30年代爆发)。不溶性物质被离心分离,得到的上层清液用于免疫沉淀反应AMPK和免疫印迹分析。gydF4y2Ba
免疫印迹和抗体gydF4y2Ba
样本煮在聚丙烯酰胺凝胶电泳样品缓冲和解决。蛋白质被转移到PVDF膜(Immobilon-FL微孔)和阻塞PBS含有5%脱脂奶粉1 h。抗体稀释在5毫升高盐渐变缓冲区(20毫米三羟甲基氨基甲烷、液pH值7.4,500毫米渐变氯化钠和0.5% (v / v)),并与细胞膜孵化。主要用于抗体免疫印迹如下:anti-AMPK-β1/2(内部)(稀释1:3,000-1:10,000吸水),anti-phospho-ACC (S79)(细胞信号,猫。不。3661),anti-phospho-AMPK (PT172)(细胞信号,猫。不。2535)anti-β-actin(σ,猫。不。A5316)。gydF4y2Ba
主要使用荧光抗体检测与二次抗体(Alexa萤石,表达载体,山羊抗兔:A21109和山羊抗小鼠:A21058)。这些都是可视化使用奥德赛红外成像仪(Li-Cor生物科学)。量化的结果是使用奥德赛执行软件和获得的信号的表示为一个比phospho-specific抗体相对合适的抗体。长篇削减免疫印迹的图片所示gydF4y2Ba补充图1gydF4y2Ba。gydF4y2Ba
醋酸纤维在下丘脑的代谢的影响gydF4y2Ba
(2 -gydF4y2Ba13gydF4y2BaC]醋酸(500毫克公斤gydF4y2Ba−1gydF4y2Ba)或(U -gydF4y2Ba13gydF4y2BaC]菊粉(100毫克)是管理8-10-week-old C57BL / 6 j雄性老鼠(查尔斯河、西班牙)通宵禁食(16 h)与i.p。(gydF4y2BangydF4y2Ba= 18)或填喂法(gydF4y2BangydF4y2Ba= 4),分别。所有老鼠和4%异氟烷麻醉(3 l / min, 99%的氧气)和脑代谢被捕1.5秒内使用大功率(5 kW)微波固定系统(tmw - 6402 c,室町Kikai有限公司,东京,日本)立即(0分),醋酸后15 - 30分钟,或菊粉180分钟后,政府。固定的大脑解剖和下丘脑和大脑切片获得和保持冷冻(−80°C),直到HR-MAS分析。gydF4y2Ba13gydF4y2BaC(125年03 MHz)gydF4y2Ba1gydF4y2BaH HR-MAS(500年,13兆赫)光谱是在11.7 T (4°C, 4000 Hz旋转)与力量皇冠宽孔光谱仪配备了MAS配件(力量Biospin, Rheinstetten,德国)。gydF4y2Ba1gydF4y2BaH-decoupledgydF4y2Ba13gydF4y2BaC HR-MAS光谱获得使用pulse-acquire序列,WALTZ-16gydF4y2Ba1gydF4y2BaH解耦应用在采集和放松的延迟时间。条件是π/ 4脉冲,8192或16384扫描([2 -gydF4y2Ba13gydF4y2Ba醋酸C]或[U -gydF4y2Ba13gydF4y2Ba分别为C]菊粉测量),64 k数据点,5 s循环延迟。的gydF4y2Ba13gydF4y2BaC光谱处理Mestrelab (gydF4y2Bahttp://mestrelab.com/gydF4y2Ba)。的化学变化gydF4y2Ba13gydF4y2BaC共振被称为醋酸C2共振(24.5件分),和gydF4y2Ba13gydF4y2BaC公司是规范化的肌醇C1 + C3共振(73.2件分),它提供了一个有用的内部参考的gydF4y2Ba13gydF4y2BaC共振可以独立规范化的组织gydF4y2Ba20.gydF4y2Ba,gydF4y2Ba47gydF4y2Ba,引入适当的修正核Overhausser增强和饱和度。gydF4y2Ba
1gydF4y2Ba从相同的活检用于H HR-MAS光谱gydF4y2Ba13gydF4y2Ba使用Carr-Purcell-Meiboom-Gil序列C HR-MAS收购是收购。采集参数5 s水pre-saturation,回波时间= 144毫秒,gydF4y2BatgydF4y2Ba= 1,女士gydF4y2BangydF4y2Ba= 100,10 kHz光谱宽度,32 k数据点和256扫描。代谢物浓度进行评估LCModel处理软件的使用修改后的版本(光谱的线性组合模型,gydF4y2Bahttp://s-provencher.com/pages/schtmgydF4y2Ba)gydF4y2Ba48gydF4y2Ba。gydF4y2Ba
使用双尾未配对的学生进行了统计检验gydF4y2BatgydF4y2Ba以及之间的值在不同的时间点或区域。观察,跌破了q1 - 1.5×差(四分位范围)或以上第三季+ 1.5×差范围(QgydF4y2Ba1gydF4y2Ba和问gydF4y2Ba3gydF4y2Ba上四分位数与下四分位数代表值和差,问的区别gydF4y2Ba3gydF4y2Ba和问gydF4y2Ba1gydF4y2Ba)被认为是异常值和不考虑统计评估。gydF4y2Ba
数据分析gydF4y2Ba
分析使用图垫棱镜(GraphPad软件、圣地亚哥、钙、美国)或占据(美国StataCorp LP,大学城,TX)。所有数据是正常使用Shapiro-Wilk测试测试。非正态分布数据日志转换为规范化的分布。对比组是由两面未配对的学生gydF4y2BatgydF4y2Ba以及对两组和单向方差分析(方差分析)gydF4y2Ba事后gydF4y2Ba图基或Bonferroni调整如果有超过两组。I.c.v.交叉注射醋酸研究使用双向成对学生的比较gydF4y2BatgydF4y2Ba以及。鉴于MEMRI和宠物的累积自然信号强度数据,信号强度剖面的差异感兴趣区域之间的所有实验群体使用哇,Mann-Whitney进行分析gydF4y2BaUgydF4y2Ba以及与商业统计软件(占据,9.1版本;StataCorp)比较概要文件全部扫描。所有结果和图形表示为±s.e.m。结果被认为是显著时gydF4y2BaPgydF4y2Ba< 0.05,显著性水平显示为*gydF4y2BaPgydF4y2Ba< 0.05,* *gydF4y2BaPgydF4y2Ba< 0.01和* * *gydF4y2BaPgydF4y2Ba< 0.001gydF4y2Ba
额外的信息gydF4y2Ba
本文引用:gydF4y2Ba霜,G。gydF4y2Baet al。gydF4y2Ba短链脂肪酸酯通过中央体内平衡机制降低食欲。gydF4y2BaCommun Nat。gydF4y2Ba5:3611 doi: 10.1038 / ncomms4611 (2014)。gydF4y2Ba
引用gydF4y2Ba
Swinburn, b . a . et al。全球肥胖流行病:由全球司机和当地环境。gydF4y2Ba《柳叶刀》gydF4y2Ba378年gydF4y2Ba,804 - 814 (2011)。gydF4y2Ba
普伦蒂斯,a . m . &杰布,美国肥胖在英国:暴食或懒惰吗?gydF4y2BaBMJgydF4y2Ba311年gydF4y2Ba,437 - 439 (1995)。gydF4y2Ba
普伦蒂斯,a . m . &杰布,美国快餐食品,能量密度和肥胖:一个可能的机械联系。gydF4y2Baob。牧师。gydF4y2Ba4gydF4y2Ba,187 - 194 (2003)。gydF4y2Ba
伊顿,s . b .祖先人类饮食:它是什么,它应该是一个范式的当代营养?gydF4y2BaProc,减轻。Soc。gydF4y2Ba65年gydF4y2Ba1 - 6 (2006)。gydF4y2Ba
Sleeth, m . L。,Thompson, E. L., Ford, H. E., Zac-Varghese, S. E. & Frost, G. Free fatty acid receptor 2 and nutrient sensing: a proposed role for fibre, fermentable carbohydrates and short-chain fatty acids in appetite regulation.减轻。牧师》。gydF4y2Ba23gydF4y2Ba,135 - 145 (2010)。gydF4y2Ba
Cani, p D。,Dewever, C. & Delzenne, N. M. Inulin-type fructans modulate gastrointestinal peptides involved in appetite regulation (glucagon-like peptide-1 and ghrelin) in rats.Br。j .减轻。gydF4y2Ba92年gydF4y2Ba,521 - 526 (2004)。gydF4y2Ba
Tolhurst, g . et al .短链脂肪酸刺激glucagon-like peptide-1通过G-protein-coupled受体FFAR2分泌。gydF4y2Ba糖尿病gydF4y2Ba61年gydF4y2Ba,364 - 371 (2012)。gydF4y2Ba
Bodinham, c . L。,Hitchen, K. L., Youngman, P. J., Frost, G. S. & Robertson, M. D. Short-term effects of whole-grain wheat on appetite and food intake in healthy adults: a pilot study.Br。j .减轻。gydF4y2Ba106年gydF4y2Ba,327 - 330 (2011)。gydF4y2Ba
Anastasovska, j . et al。可发酵碳水化合物改变下丘脑神经元活动和防止环境的致胖。gydF4y2Ba肥胖gydF4y2Ba20.gydF4y2Ba,1016 - 1023 (2012)。gydF4y2Ba
帕金森,j . r . et al .微分模式后脑干和下丘脑神经元激活的外围注入GLP-1, oxyntomodulin和氯化锂在老鼠身上被manganese-enhanced磁共振成像(MEMRI)。gydF4y2Ba科学杂志gydF4y2Ba44gydF4y2Ba,1022 - 1031 (2009)。gydF4y2Ba
Pouteau E。,Nguyen, P., Ballevre, O. & Krempf, M. Production rates and metabolism of short-chain fatty acids in the colon and whole body using stable isotopes.Proc,减轻。Soc。gydF4y2Ba62年gydF4y2Ba,87 - 93 (2003)。gydF4y2Ba
长岛,h . et al。高分辨率核磁共振光谱代谢物研究患者的脑脊液颈脊髓病、腰椎神经根病。gydF4y2Ba欧元。脊柱J。gydF4y2Ba19gydF4y2Ba,1363 - 1368 (2010)。gydF4y2Ba
Wyss, m . T。,Magistretti, P. J., Buck, A. & Weber, B. Labeled acetate as a marker of astrocytic metabolism.j . Cereb。血流金属底座。gydF4y2Ba31日gydF4y2Ba,1668 - 1674 (2011)。gydF4y2Ba
通用电气、h . et al .激活脂肪细胞的受体G protein-coupled 43导致抑制脂类分解和抑制血浆游离脂肪酸。gydF4y2Ba内分泌学gydF4y2Ba149年gydF4y2Ba,4519 - 4526 (2008)。gydF4y2Ba
肯尼,gdp等。新型多功能纳米颗粒介导核肿瘤交付、可视化和治疗肿瘤减少体内。gydF4y2Baj .控制。释放gydF4y2Ba149年gydF4y2Ba,111 - 116 (2011)。gydF4y2Ba
林,y . j . & Koretsky a . p .锰离子增强t1加权磁共振成像在脑激活:大脑功能的直接成像的方法。gydF4y2Ba粉剂。的原因。地中海。gydF4y2Ba38gydF4y2Ba,378 - 388 (1997)。gydF4y2Ba
沈,l . et al .膳食抗性淀粉增加下丘脑POMC表达式的老鼠。gydF4y2Ba肥胖(银泉)gydF4y2Ba17gydF4y2Ba降价(2009)。gydF4y2Ba
波尔多红酒,m . et al . AMPK能源是必不可少的体内平衡调节和葡萄糖传感和POMC AgRP神经元。gydF4y2Baj .中国。投资。gydF4y2Ba117年gydF4y2Ba,2325 - 2336 (2007)。gydF4y2Ba
胡,Z。,Ch一个,S. H., Chohnan, S. & Lane, M. D. Hypothalamic malonyl-CoA as a mediator of feeding behavior.Proc。《科学。美国gydF4y2BaOne hundred.gydF4y2Ba,12624 - 12629 (2003)。gydF4y2Ba
Violante, i r . et al .脑激活下丘脑的禁食诱发乳酸积累。gydF4y2Ba粉剂。的原因。地中海。gydF4y2Ba62年gydF4y2Ba,279 - 283 (2009)。gydF4y2Ba
德尔珈朵,t . C。,Violante, I. R., Nieto-Charques, L. & Cerdan, S. Neuroglial metabolic compartmentation underlying leptin deficiency in the obese ob/ob mice as detected by magnetic resonance imaging and spectroscopy methods.j . Cereb。血流金属底座。gydF4y2Ba31日gydF4y2Ba,2257 - 2266 (2011)。gydF4y2Ba
Cerdan, S。,Kunnecke, B. & Seelig, J. Cerebral metabolism of [1,2-13C2]acetate as detected by在活的有机体内gydF4y2Ba和gydF4y2Ba在体外gydF4y2Ba13 c NMR。gydF4y2Ba生物。化学。gydF4y2Ba265年gydF4y2Ba,12916 - 12926 (1990)。gydF4y2Ba
Lei, H。,Po我try-Yamate, C., Preitner, F., Thorens, B. & Gruetter, R. Neurochemical profile of the mouse hypothalamus using在活的有机体内gydF4y2Ba1 h MRS在14.1 t。gydF4y2Ba核磁共振生物医学。gydF4y2Ba23gydF4y2Ba,578 - 583 (2010)。gydF4y2Ba
因此,p . w . et al .抗性淀粉对脂肪的影响模式和中央食欲调节。gydF4y2Ba《公共科学图书馆•综合》gydF4y2Ba2gydF4y2Bae1309 (2007)。gydF4y2Ba
pswlak, d . B。,Kushner, J. A. & Ludwig, D. S. Effects of dietary glycaemic index on adiposity, glucose homoeostasis, and plasma lipids in animals.《柳叶刀》gydF4y2Ba364年gydF4y2Ba,778 - 785 (2004)。gydF4y2Ba
Darzi, J。,霜,G。S. & Robertson, M. D. Do SCFA have a role in appetite regulation?Proc,减轻。Soc。gydF4y2Ba70年gydF4y2Ba,119 - 128 (2011)。gydF4y2Ba
帕金森,j . R。,Ch一个udhri, O. B. & Bell, J. D. Imaging appetite-regulating pathways in the central nervous system using manganese-enhanced magnetic resonance imaging.神经内分泌学gydF4y2Ba89年gydF4y2Ba,121 - 130 (2009)。gydF4y2Ba
Deelchand d K。Shestov, A。人类,d . M。,Ugurbil, K. & Henry, P. G. Acetate transport and utilization in the rat brain.j . Neurochem。gydF4y2Ba109年gydF4y2Ba16(5):46 - 54岁(2009)。gydF4y2Ba
Waniewski r . a &马丁,d . l .优先利用醋酸的星形胶质细胞可归因于运输。gydF4y2Baj . >。gydF4y2Ba18gydF4y2Ba,5225 - 5233 (1998)。gydF4y2Ba
江,l . et al。增加大脑吸收和氧化醋酸的酗酒者。gydF4y2Baj .中国。投资。gydF4y2Ba123年gydF4y2Ba,1605 - 1614 (2013)。gydF4y2Ba
Hankir m K。,帕金森,j . R。,Bloom, S. R. & Bell, J. D. The effects of glutamate receptor agonists and antagonists on mouse hypothalamic and hippocampal neuronal activity shown through manganese enhanced MRI.科学杂志gydF4y2Ba59gydF4y2Ba,968 - 978 (2012)。gydF4y2Ba
哇,d . h . et al . TREK-1和Best1渠道协调快速,对GPCR激活星形胶质细胞谷氨酸释放缓慢。gydF4y2Ba细胞gydF4y2Ba151年gydF4y2Ba批准(2012)。gydF4y2Ba
迪肯,m . S。,Tooker, R. E. & Hentges, S. T. Regulation of GABA and glutamate release from proopiomelanocortin neuron terminals in intact hypothalamic networks.j . >。gydF4y2Ba32gydF4y2Ba,4042 - 4048 (2012)。gydF4y2Ba
Robertson m D。,Bickerton, A. S., Dennis, A. L., Vidal, H. & Frayn, K. N. Insulin-sensitizing effects of dietary resistant starch and effects on skeletal muscle and adipose tissue metabolism.点。j .中国。减轻。gydF4y2Ba82年gydF4y2Ba,559 - 567 (2005)。gydF4y2Ba
理查森,h。挥发性物质滥用:评估和治疗。gydF4y2Ba嗡嗡声。Toxicol。gydF4y2Ba8gydF4y2Ba,319 - 322 (1989)。gydF4y2Ba
奥谢,d . et al .神经肽Y诱导喂养的老鼠被一本小说受体介导。gydF4y2Ba内分泌学gydF4y2Ba138年gydF4y2Ba,196 - 202 (1997)。gydF4y2Ba
Paxinos, g . &华生,C。gydF4y2Ba老鼠大脑的立体定位坐标gydF4y2Ba学术出版社(2007)。gydF4y2Ba
奥利弗,m . et al . MAGfect:一种新型脂质体配方MRI标签和可视化的细胞。gydF4y2BaOrg。Biomol。化学。gydF4y2Ba4gydF4y2Ba,3489 - 3497 (2006)。gydF4y2Ba
Bhalerao s s &中央邦Harshal, a .制备、优化,表征,水杨酸脂质体稳定性的研究。gydF4y2Ba药物开发。印第安纳州。制药。gydF4y2Ba29日gydF4y2Ba,451 - 467 (2003)。gydF4y2Ba
查塔姆,j . c . &福特,j . r .乳酸和蛋白质相互作用:对NMR乳酸在生物系统的可见性。gydF4y2BaBiochim。Biophys。学报gydF4y2Ba1426年gydF4y2Ba,177 - 184 (1999)。gydF4y2Ba
赫尔希·m·G。陆,H。,Wang, T., Martin, R. J. & Baile, C. A. Intracerebroventricular (i.c.v.) administration of mouse leptin in rats: behavioral specificity and effects on meal patterns.杂志。Behav。gydF4y2Ba65年gydF4y2Ba,445 - 455 (1998)。gydF4y2Ba
争论,p . J。,Sahakian, B. J., Robbins, T. W., Koob, G. F. & Iversen, S. D. An observational method for quantifying the behavioural effects of dopamine agonists: contrasting effects of d-amphetamine and apomorphine.精神药理学(Berl)gydF4y2Ba69年gydF4y2Ba,253 - 259 (1980)。gydF4y2Ba
艾德里安,t . e . et al .人力分配和释放一种假定的新肠道激素肽YY。gydF4y2Ba胃肠病学gydF4y2Ba89年gydF4y2Ba,1070 - 1077 (1985)。gydF4y2Ba
Kreymann, b . et al .隔离和描述GLP-1 7-36酰胺从鼠肠。在糖尿病大鼠水平升高。gydF4y2Ba2月。gydF4y2Ba242年gydF4y2Ba,167 - 170 (1988)。gydF4y2Ba
贝维克·g·a . et al .小鼠hyperghrelinemia贪食的葡萄糖不能容忍,减少了瘦素的敏感性。gydF4y2Ba糖尿病gydF4y2Ba58gydF4y2Ba,840 - 846 (2009)。gydF4y2Ba
Chaudhri o . b . et al .微分下丘脑神经元激活后外围注入glp - 1在老鼠和oxyntomodulin manganese-enhanced磁共振成像探测到。gydF4y2Ba物化学。Biophys。Commun >,gydF4y2Ba350年gydF4y2Ba,298 - 306 (2006)。gydF4y2Ba
Cerdan, S。,P一个rrilla, R., Santoro, J. & Rico, M. 1H NMR detection of cerebral myo-inositol.2月。gydF4y2Ba187年gydF4y2Ba,167 - 172 (1985)。gydF4y2Ba
Provencher, s . w .估计本地化的代谢物浓度gydF4y2Ba在活的有机体内gydF4y2Ba质子核磁共振光谱。gydF4y2Ba粉剂。的原因。地中海。gydF4y2Ba30.gydF4y2Ba,672 - 679 (1993)。gydF4y2Ba
确认gydF4y2Ba
部门是由医学研究理事会(MRC)资助,生物技术和生物科学研究委员会(BBSRC),国家卫生研究所(NIHR)都来自英国,一个综合哺乳动物生物学(IMB)能力建设奖,一个FP7 -健康- 2009 - 241592之下EurOCHIP格兰特和资助NIHR帝国生物医学研究中心的资助计划。G.F. NIHR支持的高级调查员奖,A.V.由NHMRC支持海外临床研究奖学金(535976)和一个fp7格兰特-人——2009 - 256365年重返社会之下。南卡罗莱纳州和文学士saf2011赠款支持- 23622 S / BMD 23492010, ipt - 2012 - 1331 - 060000和奖学金bes - 2009 - 027615,分别。作者感谢Javier Perez先生(CSIC)专业支持在起草的插图。gydF4y2Ba
作者信息gydF4y2Ba
作者和联系gydF4y2Ba
贡献gydF4y2Ba
作者所输入的设计研究和在执行过程中扮演了一个角色的实验方法和数据收集。G.F.和大联盟协调与所有作者的手稿评论的写作。gydF4y2Ba
相应的作者gydF4y2Ba
道德声明gydF4y2Ba
相互竞争的利益gydF4y2Ba
作者声明没有竞争的经济利益。gydF4y2Ba
补充信息gydF4y2Ba
补充信息gydF4y2Ba
补充图1 (PDF 164 kb)gydF4y2Ba
权利和权限gydF4y2Ba
这项工作是基于知识共享署名3.0 Unported许可证。本文中的图片或其他第三方材料都包含在本文的创作共用许可,除非另有说明在信贷额度;如果材料不包括在Creative Commons许可下,用户需要获得许可证持有人的许可复制的材料。查看本许可证的副本,请访问http://creativecommons.org/licenses/by/3.0/gydF4y2Ba
关于这篇文章gydF4y2Ba

引用这篇文章gydF4y2Ba
霜,G。,Sleeth, M., Sahuri-Arisoylu, M.et al。gydF4y2Ba短链脂肪酸酯通过中央体内平衡机制降低食欲。gydF4y2BaNat CommungydF4y2Ba5gydF4y2Ba3611 (2014)。https://doi.org/10.1038/ncomms4611gydF4y2Ba
收到了gydF4y2Ba:gydF4y2Ba
接受gydF4y2Ba:gydF4y2Ba
发表gydF4y2Ba:gydF4y2Ba
DOIgydF4y2Ba:gydF4y2Bahttps://doi.org/10.1038/ncomms4611gydF4y2Ba
本文引用的gydF4y2Ba
一个前瞻性调查肠道微生物组成和认知能力之间的关系健康的年轻成年人gydF4y2Ba
肠道病原体gydF4y2Ba(2022)gydF4y2Ba
微生物群内不同的益生元是守恒的个人和与习惯性的纤维摄入量有关gydF4y2Ba
微生物组gydF4y2Ba(2022)gydF4y2Ba
小说的影响抗惊厥的化学Q808肠道微生物群,在pentylenetetrazole-induced癫痫大鼠海马神经递质gydF4y2Ba
BMC神经科学gydF4y2Ba(2022)gydF4y2Ba
肠脑轴在能源和葡萄糖代谢的作用gydF4y2Ba
实验与分子医学gydF4y2Ba(2022)gydF4y2Ba
猪肠道微生物群在调节宿主代谢适应寒冷的压力gydF4y2Ba
npj生物膜和微生物gydF4y2Ba(2022)gydF4y2Ba
评论gydF4y2Ba
通过提交评论你同意遵守我们的gydF4y2Ba条款gydF4y2Ba和gydF4y2Ba社区指导原则gydF4y2Ba。如果你发现一些滥用或不符合我们的条件或准则请国旗是不合适的。gydF4y2Ba
