







文摘gydF4y2Ba
严重急性呼吸系统综合症冠状病毒2 (SARS-CoV-2)已经感染了7800万人,负责超过170万人死亡。感染与变量与中和抗体水平的发展活动,可以防止感染动物模型gydF4y2Ba1gydF4y2Ba,gydF4y2Ba2 gydF4y2Ba。抗体水平下降随着时间的推移,但是,据我们所知,记忆B细胞的性质和质量,需要对再感染产生抗体还没有检查。在这里,我们报告一群87人的体液记忆反应评估感染SARS-CoV-2后在1.3和6.2个月。我们发现IgM的滴定度和免疫球蛋白抗体受体结合域(RBD)的突起蛋白SARS-CoV-2显著降低在这个时间段,IgA是影响较小。同时,等离子体中和活动减少的五倍pseudotype病毒化验。相比之下,RBD-specific记忆B细胞的数量保持不变在感染后6.2个月。记忆B细胞克隆营业额显示6.2个月后,和他们表达的抗体有更大的体细胞hypermutation,抵抗RBD突变和力量增加,表明持续进化的体液反应。免疫荧光法和PCR分析肠活检获得的无症状的个体在冠状病毒病的发病后4个月2019 (COVID-19)揭示了持久性SARS-CoV-2核酸和免疫反应性的小肠7的14个人。我们得出这样的结论:记忆B细胞反应SARS-CoV-2演化之间的1.3和6.2个月后感染的方式与抗原的持久性是一致的。gydF4y2Ba
主要gydF4y2Ba
抗体反应SARS-CoV-2最初表现为一群人没有从COVID-19约40天(1.3个月)后感染gydF4y2Ba1gydF4y2Ba。8月31日至2020年10月16日,100名参与者进行6个月随访研究访问返回。虽然最初的标准允许招生SARS-CoV-2患者的密切接触者感染经逆转录聚合酶链反应(rt - PCR)gydF4y2Ba1gydF4y2Ba13的联系人没有血清转化,被排除在进一步分析。剩余的87名参与者与RT-PCR-confirmed SARS-CoV-2感染和/或血清转化返回分析大约191天(6.2个月;165后223天)出现症状。在这个群组,症状持续了12天的中值(范围从0到44天)在急性期,和10个(11%)的参与者住院。与其他报道一致gydF4y2Ba3gydF4y2Ba,gydF4y2Ba4gydF4y2Ba38(44%)的参与者报道,持续长期症状归因于COVID-19(方法,补充表gydF4y2Ba1gydF4y2Ba,gydF4y2Ba2 gydF4y2Ba)。症状在急性疾病的持续时间和严重程度也显著大于持续急性症状在第二项研究的实验对象中访问(扩展数据图。gydF4y2Ba1 mqgydF4y2Ba)。重要的是,所有87名参与者测试为阴性SARS-CoV-2在6个月随访研究访问使用一个批准的唾液PCR试验(方法)。参与者人口和临床特点补充表所示gydF4y2Ba1gydF4y2Ba,gydF4y2Ba2 gydF4y2Ba。gydF4y2Ba
等离子体SARS-CoV-2抗体反应gydF4y2Ba
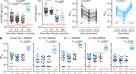
抗体反应在等离子体的RBD和核蛋白(N) SARS-CoV-2测量通过酶联免疫吸附试验(ELISA)和自动化的血清学检测gydF4y2Ba1gydF4y2Ba,gydF4y2Ba5gydF4y2Ba,gydF4y2Ba6gydF4y2Ba。Anti-RBD化验有紧密的关联(Anti-RBD免疫球蛋白ELISA和Pylon-IgG Anti-RBD IgM ELISA和Pylon-IgM 1.3个月,gydF4y2BargydF4y2Ba= 0.9200,gydF4y2BargydF4y2Ba= 0.7543,gydF4y2BaPgydF4y2Ba分别为< 0.0001)(扩展数据图。gydF4y2Ba2模拟gydF4y2Ba),anti-N化验呈中度相关(anti-N免疫球蛋白ELISA和罗氏anti-N总免疫球蛋白在1.3个月,gydF4y2BargydF4y2Ba= 0.3596,gydF4y2BaPgydF4y2Ba= 0.0012)(扩展数据图。gydF4y2Ba2 e, fgydF4y2Ba)。anti-RBD和ELISA anti-N抗体之间的等离子体明显减少(图1.3和6.2个月。gydF4y2Ba1模拟gydF4y2Ba)。值得注意的是,降低绑定活动实质上不同的同形像和目标。IgM显示anti-RBD反应最大的减少(53%),其次是免疫球蛋白(32%);anti-RBD IgA只下降了15%,anti-N IgG水平22%(图。gydF4y2Ba1 egydF4y2Ba)。相比之下,罗氏anti-N化验gydF4y2Ba6gydF4y2Ba显示一个小但显著增加(19%)在两个时间点之间的反应性(扩展数据图。gydF4y2Ba2 ggydF4y2Ba),这可能解释为抗原的使用桥接方法gydF4y2Ba7gydF4y2Ba。在所有情况下,减少的大小成反比,与初始抗体水平直接相关,这样初始水平较高的个体,显示更大的相对变化(图。gydF4y2Ba外:我1gydF4y2Ba,扩展数据图。gydF4y2Ba2 hgydF4y2Ba)。两个时间点之间的所有测量有紧密的关联(扩展数据图。gydF4y2Ba2我gydF4y2Ba)和anti-N免疫球蛋白与各自的IgM,免疫球蛋白和IgA anti-RBD反应在1.3个月(扩展数据图。gydF4y2Ba2阻燃剂gydF4y2Ba)。值得注意的是,患有急性症状持续的水平明显高于有anti-RBD免疫球蛋白和anti-N总抗体在研究访问(扩展数据图。gydF4y2Ba1 lgydF4y2Ba)。gydF4y2Ba

一个gydF4y2Ba- - - - - -gydF4y2BadgydF4y2Baelisa结果测量等离子体反应RBD (gydF4y2Ba一个gydF4y2Ba,gydF4y2BabgydF4y2Ba,gydF4y2BacgydF4y2Ba)和N蛋白(gydF4y2BadgydF4y2Ba)在初始1.3——6.2月复诊,分别。gydF4y2Ba一个gydF4y2Ba,Anti-RBD IgM。gydF4y2BabgydF4y2Ba,Anti-RBD免疫球蛋白。gydF4y2BacgydF4y2Ba,Anti-RBD IgAgydF4y2BadgydF4y2Ba,Anti-N免疫球蛋白。归一化曲线下的面积(AUC)值87人所示gydF4y2Ba一个gydF4y2Ba- - - - - -gydF4y2BadgydF4y2Ba两个时间点。积极的和消极的控制包括进行验证gydF4y2Ba1gydF4y2Ba。gydF4y2BaegydF4y2Ba等离子体之间的抗体水平、相对变化anti-RBD IgM 1.3和6.2个月,免疫球蛋白,IgA anti-N免疫球蛋白在所有87人。gydF4y2BafgydF4y2Ba- - - - - -gydF4y2Ba我gydF4y2Ba相对变化量之间的抗体水平1.3和6.2个月策划与相应的抗体水平在1.3个月。gydF4y2BafgydF4y2Ba,Anti-RBD IgM。gydF4y2BargydF4y2Ba=−0.83,gydF4y2BaPgydF4y2Ba< 0.0001。gydF4y2BaggydF4y2Ba,Anti-RBD免疫球蛋白。gydF4y2BargydF4y2Ba=−0.76,gydF4y2BaPgydF4y2Ba< 0.0001。gydF4y2BahgydF4y2Ba,Anti-RBD IgA。gydF4y2BargydF4y2Ba=−0.67,gydF4y2BaPgydF4y2Ba< 0.0001。gydF4y2Ba我gydF4y2Ba,Anti-N免疫球蛋白。gydF4y2BargydF4y2Ba=−0.87,gydF4y2BaPgydF4y2Ba< 0.0001。gydF4y2BajgydF4y2Ba,排名平均NTgydF4y2Ba50gydF4y2Ba在1.3个月(蓝色)和6.2个月(红色)的87人进行了研究。gydF4y2BakgydF4y2Ba图表显示,NTgydF4y2Ba50gydF4y2Ba血浆从所有87人收集在1.3和6.2个月。gydF4y2BaPgydF4y2Ba< 0.0001。gydF4y2BalgydF4y2Ba相对变化量之间的等离子体中和滴定度1.3和6.2个月策划相应的滴定度在1.3个月。为gydF4y2Ba一个gydF4y2Ba- - - - - -gydF4y2BaegydF4y2Ba,gydF4y2BakgydF4y2Ba绘制的价值观和单杠表示几何平均数。统计学意义是决定使用双尾Wilcoxon配对符号秩检验gydF4y2Ba一个gydF4y2Ba- - - - - -gydF4y2BadgydF4y2Ba,gydF4y2BakgydF4y2Ba邓恩,弗里德曼的多重比较检验gydF4y2BaegydF4y2Ba。的gydF4y2BargydF4y2Ba和gydF4y2BaPgydF4y2Ba值gydF4y2BafgydF4y2Ba- - - - - -gydF4y2Ba我gydF4y2Ba,gydF4y2BalgydF4y2Ba是由双尾枪兵的相关性。gydF4y2Ba
我们测量等离子体中和活动使用的hiv - 1病毒准型SARS-CoV-2峰值蛋白质gydF4y2Ba1gydF4y2Ba,gydF4y2Ba8gydF4y2Ba。与其他报道一致gydF4y2Ba9gydF4y2Ba,gydF4y2Ba10gydF4y2Ba,几何平均数half-maximal中和滴定度(NTgydF4y2Ba50gydF4y2Ba)这组87名参与者在1.3和6.2个月,401年和78年respectively-representing降低5倍(无花果。gydF4y2Ba1 j, kgydF4y2Ba)。中和活动直接与免疫球蛋白anti-RBD ELISA测量(扩展数据图。gydF4y2Ba2 q, rgydF4y2Ba)。此外,减少中和活动的绝对星等是成反比,直接与中和活动在更早的时间点(图。gydF4y2Ba1 lgydF4y2Ba)。我们得出这样的结论:抗体RBD和等离子体中和活动显著减少,但仍可检测,6个月后感染SARS-CoV-2大多数人。gydF4y2Ba
检查循环B细胞的表型景观,我们进行高维流式细胞术在41个随机选择的个体在时间点和比较结果pre-COVID-19健康个体样本(gydF4y2BangydF4y2Ba= 20)。全球高维映射gydF4y2BatgydF4y2Ba分布式随机邻居嵌入显示显著持久改变的人从COVID-19(扩展数据图中恢复过来。gydF4y2Ba3gydF4y2Ba)。集群的相对表现2、7、8和10(对应于天真、内存plasmablast和浆细胞,分别)降低为1.3个月,所以在以后的时间点。Metacluster 15对应于不成熟B细胞从骨头含有新移民在早期的时间点增加,但回到控制水平的观察期间(扩展数据图。gydF4y2Ba3罪犯gydF4y2Ba)。gydF4y2Ba
SARS-CoV-2记忆B细胞剧目gydF4y2Ba
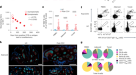
而浆细胞循环抗体的来源,记忆B细胞导致召回响应。识别和枚举循环SARS-CoV-2-specific记忆B细胞室,我们使用流式细胞仪分离单个B淋巴细胞必将RBD的受体gydF4y2Ba1gydF4y2Ba(无花果。gydF4y2Ba2gydF4y2Ba,扩展数据图。gydF4y2Ba4gydF4y2Ba)。值得注意的是,RBD-binding记忆B细胞的比例略微增加1.3和6.2个月21之间随机选择的个体(图。gydF4y2Ba2 bgydF4y2Ba)。gydF4y2Ba

一个gydF4y2Ba流式细胞术,代表情节展示双重alexafluor - 647 - rbd和PE-RBD-binding B细胞6研究个体(指定COV21, COV47 COV57, COV72, COV96和COV107)(控制策略扩展数据图所示。gydF4y2Ba5gydF4y2Ba)。抗原B细胞的百分比表示。gydF4y2BabgydF4y2Ba,如gydF4y2Ba一个gydF4y2Ba。图总结RBD-binding记忆B细胞样本中获得的比例在1.3和6.2个月来自21个随机选择的个体。红色单杠显示几何平均值。统计学意义是决定使用双尾Wilcoxon配对符号秩检验。gydF4y2BacgydF4y2Ba、体细胞数核苷酸突变IGVH(上)和IGVL(底部)基因抗体获得1.3或6.2个月后从指定的个人(左)和六个捐助者(右)。使用双尾Mann-Whitney确定统计学意义gydF4y2BaUgydF4y2Ba测试。单杠表示中位数的值。gydF4y2BadgydF4y2Ba饼图显示,抗体的分布序列从后六个人1.3gydF4y2Ba1gydF4y2Ba(上)或6.2个月(底部)。内部圈子中的数字显示的数量为上面所示的单个圆序列分析。饼图分区大小正比于无性生殖相关序列的数量。无性繁殖系地扩大的黑色轮廓表示频率序列中发现每个参与者。彩色切片表明坚持克隆(相同IGHV和IGLV基因和高度类似CDR3上)发现在这两个时间点相同的参与者。灰色片表明克隆的时间点。白片显示背心发现这两个时间点,剩下的白色区域表明序列被孤立的一次。gydF4y2BaegydF4y2Ba图表显示,在两个时间点相对单克隆所有六个捐助者。红色单杠表示平均值。统计学意义是决定使用双尾gydF4y2BatgydF4y2Ba以及。gydF4y2Ba
来确定是否有改变记忆B细胞产生的抗体6.2个月后,我们得到532配对抗体重型和轻型链相同的6人检查在更早的时间点gydF4y2Ba1gydF4y2Ba(补充表gydF4y2Ba3gydF4y2Ba)。没有显著差异的代表性IGV基因在两个时间点,其中的代表gydF4y2BaIGHV3-30gydF4y2Ba和gydF4y2BaIGHV3-53gydF4y2Ba基因片段gydF4y2Ba1gydF4y2Ba,gydF4y2Ba11gydF4y2Ba,gydF4y2Ba12gydF4y2Ba,gydF4y2Ba13gydF4y2Ba,gydF4y2Ba14gydF4y2Ba,gydF4y2Ba15gydF4y2Ba,gydF4y2Ba16gydF4y2Ba(扩展数据图。gydF4y2Ba5 a、bgydF4y2Ba)。符合这一观点(类似于前面的时间点),抗体,共享同一IGHV IGLV基因占8.6%的序列在不同的个体(扩展数据图。gydF4y2Ba5度gydF4y2Ba)。正如所预期的那样,有一个小——但有限明显改善的百分比增加IgG-expressing anti-RBD记忆细胞,从49%到58% (gydF4y2BaPgydF4y2Ba= 0.011)(扩展数据图。gydF4y2Ba5 d-fgydF4y2Ba)。符合分数提高免疫球蛋白g记忆细胞,体细胞的程度hypermutation本和IGL在两个时间点之间的所有六个人。而核苷酸突变的平均数量在本和IGL只有4.2和2.8,分别在第一个时间点,这些值增加到11.7和6.5,分别在第二个时间点(gydF4y2BaPgydF4y2Ba< 0.0001)(图gydF4y2Ba2摄氏度gydF4y2Ba,扩展数据图。gydF4y2Ba6 fgydF4y2Ba)。相比之下,整体的平均长度和疏水性complementarity-determining地区3 (CDR3上)的本和IGL持平(扩展数据图。gydF4y2Ba6克,hgydF4y2Ba)。gydF4y2Ba
类似于前面的时间点,我们发现扩展记忆B细胞的克隆6.2个月(包括23个克隆,出现在两个时间点)。然而,扩大克隆只占12.4%所有抗体序列后6.2个月,而32%后1.3个月(gydF4y2BaPgydF4y2Ba= 0.0225)(图gydF4y2Ba2 d, egydF4y2Ba)。此外,内存间的整体克隆组成不同的两个时间点的所有个人我们检查(无花果。gydF4y2Ba二维gydF4y2Ba)。43克隆扩张,出现在更早的时间点没有检测到6.2个月后,和22个新的扩展克隆出现了。此外,克隆的相对分布,出现在两个时间点也不同。例如,占主导地位的个人指定COV21克隆和COV57-in他们代表所有序列的9.0%和16.7%,分别为1.3个月后都是减少到1.1%和1.9%,分别后的序列(图6.2个月。gydF4y2Ba二维gydF4y2Ba补充表gydF4y2Ba3gydF4y2Ba)。我们的结论是,虽然RBD-specific记忆B细胞的大小之间的隔间是守恒的感染SARS-CoV-2 1.3和6.2个月后,有大量的克隆营业额和抗体序列进化与长时间的生发中心反应是一致的。gydF4y2Ba
SARS-CoV-2单克隆抗体gydF4y2Ba
我们测试了122名代表抗体6.2月时间点反应RBD(补充表gydF4y2Ba4gydF4y2Ba)。抗体,我们评估包括:(1)49抗体被随机选择从这些抗体只出现一次;(2)23抗体出现单打在1.3和6.2个月;(3)23代表新出现的克隆扩张;和(4)27日扩大克隆的代表出现在两个时间点。一百一十五的122 RBD结合抗体,这表明流式细胞术有效地识别B细胞产生抗体anti-RBD(无花果。gydF4y2Ba3gydF4y2Ba补充表gydF4y2Ba4gydF4y2Ba,gydF4y2Ba5gydF4y2Ba)。抗体在一起,意味着ELISA half-maximal有效浓度(ECgydF4y2Ba50gydF4y2Ba)没有明显不同的两个时间点gydF4y2Ba1gydF4y2Ba(无花果。gydF4y2Ba3gydF4y2Ba补充表gydF4y2Ba4gydF4y2Ba)。然而,比较的抗体出现在两个时间点显示EC的显著改善gydF4y2Ba50gydF4y2Ba6.2个月后(gydF4y2BaPgydF4y2Ba= 0.0227)(图gydF4y2Ba3 bgydF4y2Ba,扩展数据图。gydF4y2Ba7一个gydF4y2Ba)。gydF4y2Ba

一个gydF4y2Ba图表显示,anti-SARS-CoV-2 RBD抗体反应。ELISA ECgydF4y2Ba50gydF4y2Ba值为所有抗体以1.3个月gydF4y2Ba1gydF4y2Ba,122人选择单克隆抗体以6.2个月。单杠表示几何平均数。使用双尾Mann-Whitney确定统计学意义gydF4y2BaUgydF4y2Ba以及。gydF4y2BabgydF4y2Ba,电子商务gydF4y2Ba50gydF4y2Ba值为所有52抗体出现在1.3和6.2个月。两个或两个以上的实验的平均水平。单杠表示几何平均数。统计学意义是决定使用双尾Wilcoxon配对符号秩检验。gydF4y2BacgydF4y2Ba,RBD的表面表示ACE2-binding足迹显示虚线和选定的残留物中发现流行株(灰色)和调节抵抗力的残留二班(红色)(C144)和3(绿色)(C135)抗体了。gydF4y2BadgydF4y2Ba图表显示,ELISA绑定曲线C144(黑色虚线)及其克隆获得的亲戚后6.2个月(C050、C051 C052, C053和C054)(实线)绑定到野生型(WT) Q493R, R346S E484K rbd。gydF4y2BaegydF4y2Ba。热图显示日志gydF4y2Ba2 gydF4y2Ba改变相对折叠EC的变化gydF4y2Ba50gydF4y2Ba对表示RBD突变体26抗体克隆对获得1.3和6.2个月的变化反应最为显著。每个抗体的原产地参与者对上面的指示。所有的实验都至少执行两次。gydF4y2Ba
是否表达的抗体记忆B细胞在后期时间点还显示宽度改变,我们比他们早克隆亲戚绑定使用控制和突变rbd化验。替换E484K Q493RgydF4y2Ba17gydF4y2Ba被选为抗二班抗体(如C144和C121)直接绑定到RBD ACE2-interaction岭gydF4y2Ba1gydF4y2Ba,gydF4y2Ba18gydF4y2Ba,gydF4y2Ba19gydF4y2Ba,gydF4y2Ba20.gydF4y2Ba,R346S N439K, N440K选择抗3班抗体(如C135)不直接干涉ACE2绑定gydF4y2Ba1gydF4y2Ba,gydF4y2Ba17gydF4y2Ba,gydF4y2Ba18gydF4y2Ba,gydF4y2Ba19gydF4y2Ba,gydF4y2Ba20.gydF4y2Ba(无花果。gydF4y2Ba3 cgydF4y2Ba)。此外,我们还测试了V367F A475V, S477N和V483A RBD的突变体,代表循环变异赋予完全或部分抵抗1级和2抗体gydF4y2Ba17gydF4y2Ba,gydF4y2Ba18gydF4y2Ba,gydF4y2Ba21gydF4y2Ba(无花果。gydF4y2Ba3 cgydF4y2Ba)。52的抗体克隆对出现在这两个时间点,43(83%)显示总体增加绑定在6.2月时间点突变rbd(扩展数据图。gydF4y2Ba7常数gydF4y2Ba补充表gydF4y2Ba5gydF4y2Ba)。例如,C144(抗体在1.3个月的时间恢复点)无法绑定到RBD (Q493R)或RBD (E484K),但是它的所有五个克隆衍生品收集在6.2个月必定RBD (Q493R)和一个还显示绑定RBD (E484K)(图。gydF4y2Ba3 dgydF4y2Ba)。总的来说,最为明显增加绑定发生突变影响RBD氨基酸位置如E484 Q493, N439, N440 R346,二班和3抗体的结合的关键gydF4y2Ba17gydF4y2Ba,gydF4y2Ba18gydF4y2Ba(无花果。gydF4y2Ba3 egydF4y2Ba,扩展数据图。gydF4y2Ba7常数gydF4y2Ba补充表gydF4y2Ba5gydF4y2Ba)。gydF4y2Ba
接下来,我们测试了所有122抗体6.2月为活动时间点准型SARS-CoV-2中和试验gydF4y2Ba1gydF4y2Ba,gydF4y2Ba8gydF4y2Ba(无花果。gydF4y2Ba4gydF4y2Ba补充表gydF4y2Ba6gydF4y2Ba)。符合RBD-binding化验,意味着中和half-maximal抑制浓度(ICgydF4y2Ba50gydF4y2Ba)值没有显著不同的在两个时间点当所有抗体进行比较gydF4y2Ba1gydF4y2Ba(无花果。gydF4y2Ba4gydF4y2Ba)。然而,比较的抗体出现在两个时间点显示的显著改善集成电路gydF4y2Ba50gydF4y2Ba值为6.2个月(gydF4y2BaPgydF4y2Ba= 0.0003)(图gydF4y2Ba4 bgydF4y2Ba,扩展数据图。gydF4y2Ba8gydF4y2Ba)。gydF4y2Ba

一个gydF4y2Ba,SARS-CoV-2 pseudovirus中和试验。集成电路gydF4y2Ba50gydF4y2Ba值为所有抗体以1.3个月gydF4y2Ba1gydF4y2Ba,122人选择抗体以6.2个月。抗体与集成电路gydF4y2Ba50gydF4y2Ba值高于1μg毫升gydF4y2Ba−1gydF4y2Ba在绘制1μg毫升gydF4y2Ba−1gydF4y2Ba。意思是两个独立的实验。红酒吧表示几何平均数。使用双尾Mann-Whitney确定统计学意义gydF4y2BaUgydF4y2Ba以及。gydF4y2BabgydF4y2Ba,集成电路gydF4y2Ba50gydF4y2Ba值52抗体出现在1.3和6.2个月。红酒吧表示几何平均数。统计学意义是决定使用双尾Wilcoxon配对符号秩检验。gydF4y2BacgydF4y2Ba,集成电路gydF4y2Ba50gydF4y2Ba值5双单克隆抗体克隆亲属获得1.3或6.2个月后中和的野生型和突变体SARS-CoV-2 pseudovirus。抗体标识符的1.3月- 6.2月单克隆抗体对表示。大胆的造型表示与大幅增加中和抗体对活动后6.2个月。gydF4y2BadgydF4y2Ba,图显示了归一化相对发光单元(RLU)细胞溶解产物293 t细胞表达ACE2,感染后48 h SARS-CoV-2 pseudovirus包含野生型RBD或三种突变RBD (Q493R, E484G和R346S)的两种单克隆抗体浓度增加C144(1.3个月)(虚线)或C051(6.2个月)(实线)。实验至少进行两次。gydF4y2BaegydF4y2Ba,C051绑定模型。两个相邻的表面表示“下”RBD (RBD)gydF4y2Ba一个gydF4y2Ba和RBDgydF4y2BaBgydF4y2Ba)三聚物,与C144抗原决定基在青色rbd强调和立场的氨基酸突变积累到父抗体相比,C051 C144强调把侧链在CαC051 V的atom表示gydF4y2BaHgydF4y2BaVgydF4y2BalgydF4y2Ba绑定到邻rbd。C051交互有两个rbd是建模的基础上低温电子显微镜C144工厂一定会飙升三聚物的结构gydF4y2Ba18gydF4y2Ba。HC,重链;LC,轻链。gydF4y2Ba
确定抗体出现改变RBD绑定也显示中和宽度增加,我们测试了5个代表抗体对恢复在两个时间点对hiv - 1病毒准型与E484G Q493R, R346S突变蛋白质(图。gydF4y2Ba4摄氏度gydF4y2Ba补充表gydF4y2Ba6gydF4y2Ba)。值得注意的是,Q493R和E484G准型病毒被C144耐中和;相比之下,C051(6.2月克隆的导数C144)中和两种变体,与集成电路gydF4y2Ba50gydF4y2Ba4.7和3.1 ng毫升的价值观gydF4y2Ba−1gydF4y2Ba分别(无花果。gydF4y2Ba4 c, dgydF4y2Ba)。同样,R346S准型病毒抵抗C032,但C080(6.2月克隆的导数C032)中和这种变体ICgydF4y2Ba50gydF4y2Ba5.3 ng毫升gydF4y2Ba−1gydF4y2Ba(无花果。gydF4y2Ba4摄氏度gydF4y2Ba,扩展数据图。gydF4y2Ba8 b-fgydF4y2Ba)。绑定和中和活动与观察到的变化一致,几个late-appearing抗体(例如,C051)中直接获得的突变或相邻RBD-binding抗体结合部位(图gydF4y2Ba4 egydF4y2Ba,扩展数据图。gydF4y2Ba8 g-jgydF4y2Ba)。我们得出这样的结论:记忆B细胞进化在观察期间表达与中和效力和广度增加抗体。gydF4y2Ba
SARS-CoV-2抗原持久性gydF4y2Ba
抗体进化发生在体细胞突变和选择在生发中心抗原可以保留免疫复合物的形式表面上的滤泡树突细胞的时间太长。剩余的蛋白质在组织代表另一个潜在的抗原来源。SARS-CoV-2在ACE2-expressing复制细胞在肺部,鼻咽和小肠gydF4y2Ba22gydF4y2Ba,gydF4y2Ba23gydF4y2Ba,gydF4y2Ba24gydF4y2Ba,gydF4y2Ba25gydF4y2Ba,病毒RNA在粪便样本中发现即使在鼻咽的病毒清除gydF4y2Ba26gydF4y2Ba,gydF4y2Ba27gydF4y2Ba,gydF4y2Ba28gydF4y2Ba。以确定是否有可能在肠道抗原持久性解决临床疾病后,我们获得了上、下消化道的活检14个人平均4个月(范围2.8 ~ 5.7个月)后初始COVID-19诊断(补充表gydF4y2Ba7gydF4y2Ba)。疣状进行,以确定病毒蛋白也可检测上下消化道,从个人可以消除识别信息活检大流行(gydF4y2BangydF4y2Ba= 10)作为控制。ACE2 SARS-CoV-2 N蛋白肠肠上皮细胞中检测到5(图14个人。gydF4y2Ba5模拟gydF4y2Ba无花果、扩展数据。gydF4y2Ba9gydF4y2Baa -gydF4y2Ba10 a, bgydF4y2Ba补充表gydF4y2Ba7gydF4y2Ba),但无法控制样品(扩展数据图。gydF4y2Ba9我gydF4y2Ba)。当检测到,疣状是零星的,不完整的,独家的肠上皮细胞与炎性浸润无关(扩展数据无花果。gydF4y2Ba9gydF4y2Baa -gydF4y2Ba10 a, bgydF4y2Ba)。临床批准nasopharyngeal-swab PCR检测为阴性的14个人的活组织检查。然而,活检样本3的14个参与者产生了PCR扩增子sequence-verified SARS-CoV-2(方法,补充表gydF4y2Ba7gydF4y2Ba)。此外,病毒RNA原位杂交检测的活检样本两个参与者测试(扩展数据图。gydF4y2Ba10 c, dgydF4y2Ba),但无法控制样品(扩展数据图。gydF4y2Ba10 egydF4y2Ba)。抽样可变性在内窥镜手术可能导致不完整的病毒RNA和蛋白质之间的一致性检测化验。gydF4y2Ba

一个gydF4y2BaEPCAM人类肠上皮细胞,免疫荧光图像彩色(红色),DAPI(蓝色)和ACE2(绿色gydF4y2Ba一个gydF4y2Ba,gydF4y2BacgydF4y2Ba)或SARS-CoV-2 N(绿色gydF4y2BabgydF4y2Ba,gydF4y2BadgydF4y2Ba)在肠活检92 d COVID-19症状发作后参与者cgi - 088,在末端回肠(gydF4y2Ba一个gydF4y2Ba,gydF4y2BabgydF4y2Ba)或十二指肠(gydF4y2BacgydF4y2Ba,gydF4y2BadgydF4y2Ba)。地区白盒子正确的小组gydF4y2Ba一个gydF4y2Ba,gydF4y2BacgydF4y2Ba扩大所示gydF4y2BabgydF4y2Ba,gydF4y2BadgydF4y2Ba,分别。箭头表示可检测到SARS-CoV-2肠上皮细胞抗原。酒吧,规模100μm。至少两次独立实验重复类似的结果。gydF4y2Ba
中和抗体SARS-CoV-2发展大多数人感染后但随时间衰减gydF4y2Ba7gydF4y2Ba,gydF4y2Ba9gydF4y2Ba,gydF4y2Ba10gydF4y2Ba,gydF4y2Ba29日gydF4y2Ba,gydF4y2Ba30.gydF4y2Ba,gydF4y2Ba31日gydF4y2Ba,gydF4y2Ba32gydF4y2Ba。这些抗体是有效的预防和治疗在动物模型中,可能会有一个角色在防止再感染人类gydF4y2Ba2 gydF4y2Ba。虽然有显著降低血浆中和活动1.3和6.2个月,抗体滴定度保持在大多数个人可衡量的gydF4y2Ba9gydF4y2Ba,gydF4y2Ba10gydF4y2Ba,gydF4y2Ba29日gydF4y2Ba,gydF4y2Ba30.gydF4y2Ba,gydF4y2Ba31日gydF4y2Ba,gydF4y2Ba32gydF4y2Ba,gydF4y2Ba33gydF4y2Ba,gydF4y2Ba34gydF4y2Ba,gydF4y2Ba35gydF4y2Ba,gydF4y2Ba36gydF4y2Ba。gydF4y2Ba
单克隆抗体中和从个人获得早期康复期间显示明显的低水平的体细胞突变,一些先前的报道归因于生发中心形成缺陷gydF4y2Ba11gydF4y2Ba,gydF4y2Ba12gydF4y2Ba,gydF4y2Ba14gydF4y2Ba,gydF4y2Ba37gydF4y2Ba,gydF4y2Ba38gydF4y2Ba,gydF4y2Ba39gydF4y2Ba,gydF4y2Ba40gydF4y2Ba。我们的数据表明,anti-SARS-CoV-2记忆B细胞反应的发展在前六个月后感染,免疫球蛋白体细胞突变的积累,生产的抗体中和广度和强度增加。持续的抗体进化发生在生发中心和要求B细胞暴露于抗原被困在滤泡树突细胞免疫复合物的形式gydF4y2Ba41gydF4y2Ba。这种形式的抗原可以长寿,因为滤泡树突细胞免疫复合物内化。此外,即使少量的持久病毒抗原抗体进化可能加剧。的观察SARS-CoV-2信使rna和蛋白质在小肠上皮细胞仍可检测一些人在几个月后感染是一致的相对持久anti-RBD IgA抗体和抗体持续进化gydF4y2Ba34gydF4y2Ba,gydF4y2Ba35gydF4y2Ba,gydF4y2Ba36gydF4y2Ba。gydF4y2Ba
记忆反应负责防止再感染,为有效的疫苗接种是必不可少的。观察记忆B细胞反应不衰变后6.2个月gydF4y2Ba34gydF4y2Ba,gydF4y2Ba35gydF4y2Ba,gydF4y2Ba42gydF4y2Ba,而是继续发展,强烈暗示,人感染SARS-CoV-2可能迅速有效地应对病毒再次接触。gydF4y2Ba
方法gydF4y2Ba
数据报告gydF4y2Ba
没有统计方法被用来预先确定样本容量。实验并不随机,调查人员没有被分配在实验和结果评估。gydF4y2Ba
研究参与者gydF4y2Ba
以前注册研究的参与者gydF4y2Ba1gydF4y2Ba被要求换取6个月复诊洛克菲勒大学医院从8月31日到2020年10月16日。合格受试者18 - 76岁的成年人都患有SARS-CoV-2 rt - pcr(例)或感染的密切接触者(例如,同一个家庭的成员,同事或相同的宗教团体的成员)的人被诊断出患有SARS-CoV-2感染通过rt - pcr(联系人)。没有密切接触者血清转化对SARS-CoV-2评估血清学检测(“高通量自动化血清学检测”中描述)并不包括在后续的分析。大多数研究参与者被大纽约地区的三个州的居民被要求返回大约6个月后COVID-19症状发作的时间。参与者提出了洛克菲勒大学医院的血液样本收集和被要求召回在急性症状和临床表现的严重程度(前六周)和恢复期的直到第二次研究访问(7周)COVID-19阶段,分别。急性感染的严重程度评估由世界卫生组织(世卫组织)“序数临床进展/改进规模”(gydF4y2Bahttps://www.who.int/publications/i/item/covid-19-therapeutic-trial-synopsisgydF4y2Ba)。呼吸急促是通过修改后的医学研究委员会评估呼吸困难gydF4y2Ba43gydF4y2Ba。参与者提供持续的症状归因于COVID-19被确定的基础上慢性呼吸急促或疲劳,赤字运动能力和/或三个或更多额外的长期持久的不明原因的发烧等症状,胸痛、最近诊断为心脏sequalae,关节痛,损伤的浓度或精神敏锐,嗅觉或味觉障碍,神经病变或皮肤的发现gydF4y2Ba3gydF4y2Ba,gydF4y2Ba4gydF4y2Ba。所有参与者在洛克菲勒大学提供书面知情同意参与这项研究,研究符合良好的临床实践。临床数据收集和管理被iMedRIS使用软件进行虹膜。这项研究是符合所有相关的伦理规定和执行协议的研究与人类参与者的机构审查委员会批准了洛克菲勒大学(IRB)。gydF4y2Ba
胃肠检查队列gydF4y2Ba
确定SARS-CoV-2可以存在于胃肠道,我们招募了一群14个人,之前从COVID-19疾病诊断和恢复。合格的参与者包括成人,18 - 76岁的曾诊断为SARS-CoV-2通过rt - pcr或结合临床症状符合COVID-19 +血清转化的证据,并呈现给西奈山医院的胃肠病学诊所。内窥镜手术进行临床表示条件的详细补充表gydF4y2Ba7gydF4y2Ba。所有与会者都无症状时内镜程序和阴性SARS-CoV-2 nasal-swab PCR(循环阈值(gydF4y2BaCgydF4y2BatgydF4y2Ba)截止< 38)。gydF4y2Ba
西奈山的CLIA-certified实验室卫生系统验证了laboratory-developed nasopharyngeal-swab实时rt - pcr测试根据纽约州卫生部Wadsworth SARS-CoV-2中心验证过程gydF4y2Ba44gydF4y2Ba。从所有参与者获得知情同意。biopsy-related研究西奈山伦理委员会批准/ IRB (IRB 16 - 0583,“病毒感染的影响及其治疗胃肠道免疫细胞”)。gydF4y2Ba
SARS-CoV-2唾液PCR测试gydF4y2Ba
SARS-CoV-2 PCR方法对唾液样本开发及其性能特征由洛克菲勒大学临床基因组学实验室。这laboratory-developed测试已经被纽约州授权在紧急情况下使用授权供授权实验室使用。唾液收集到硫氰酸胍缓冲(如前所述)gydF4y2Ba45gydF4y2Ba。RNA提取使用一个基于列(试剂盒QIAmp DSP病毒RNA迷你包,61904)或magnetic-bead-based方法如前所述gydF4y2Ba46gydF4y2Ba。Reverse-transcribed cDNA是放大使用引物和探针验证由疾病控制与预防中心或哥伦比亚大学个性化药物基因组学实验室,分别经美国食品和药物管理局批准下紧急使用授权。病毒RNA被认为是发现如果gydF4y2BaCgydF4y2BatgydF4y2Ba两个病毒引物和探针组合是< 40岁。gydF4y2Ba
血液样本处理和存储gydF4y2Ba
外周血单核细胞是通过梯度离心法和储存在液氮在FCS的存在和DMSO溶液。肝素化血浆和血清样本被整除,储存在−20°C以下。在实验之前,整除的血浆样本heat-inactivated (56°C 1 h)然后储存在4°C。gydF4y2Ba
高通量自动化血清学检测gydF4y2Ba
血浆样品从80年的87名参与者被高通量测试自动化的血清学检测。罗氏的Elecsys anti-SARS-CoV-2试验进行罗氏Cobas e411(罗氏诊断)。Elecsys反检测SARS所致CoV-2试验使用一个代表N的重组蛋白抗原抗体的测定SARS所致浸2。这个试验收到紧急使用授权美国食品和药物管理局的批准gydF4y2Ba6gydF4y2Ba。的桥塔COVID-19 IgG、IgM化验是用来测量血浆免疫球蛋白和SARS-CoV-2 IgM抗体,分别。血浆样本化验在塔3 d分析器(ET医疗)如前所述gydF4y2Ba5gydF4y2Ba。这个临床试验实施作为laboratory-developed测试在纽约州卫生部规定。总之,使用组合试验进行测试条包含井predispensed试剂。COVID-19试剂包含生物素化的重组版本的SARS-CoV-2 S-protein RBD和微量的N蛋白作为抗原,免疫球蛋白结合IgM,分别。截止值为塔化验测定使用的均值non-COVID-19南达科他州+ 6样品。样品的结果报告的形式截止索引或索引值,这取决于测试的仪器读出样品除以仪器读出截止。gydF4y2Ba
elisagydF4y2Ba
elisagydF4y2Ba47gydF4y2Ba,gydF4y2Ba48gydF4y2Ba评估抗体绑定SARS-CoV-2 N (40588 - v08b嘉汉生物),RBD和额外的RBD的涂层进行high-binding 96 -半孔板(康宁3690)50μl / 1μg毫升gydF4y2Ba−1gydF4y2Ba蛋白质溶液在磷酸盐(PBS)一夜之间在4°C。洗盘子都洗了6次缓冲区(1×PBS Tween-20 0.05% (Sigma-Aldrich))和孵化170μl /阻断缓冲区(1×PBS Tween-20 BSA为2%和0.05%(σ))在室温下1 h。后立即阻止,单克隆抗体或血浆样本中添加PBS和孵化1 h在室温下。血浆样本化验1:67开始稀释和7系列稀释额外的3倍。单克隆抗体测试10μg毫升gydF4y2Ba−1gydF4y2Ba开始连续稀释浓度和10个额外的4倍。洗盘子洗了6次,缓冲然后孵化反人类免疫球蛋白,IgM或IgA二级抗体结合辣根过氧化物酶(合)(杰克逊免疫研究109-036-088 109-035-129σA0295)在阻断缓冲区1:5,000稀释(IgM和免疫球蛋白)或1:3,000稀释(IgA)。板是由合的衬底,三甲(ThermoFisher) 10分钟(血浆样本)或4分钟(单克隆抗体),然后发展中反应是停止通过添加50μl 1 M HgydF4y2Ba2 gydF4y2Ba所以gydF4y2Ba4gydF4y2Ba和吸光度测量在450 nm ELISA标(FluoStarω,BMG Labtech)与ω和ω火星软件进行分析。血浆样本,积极控制(等离子体从参与者COV72稀释66.6倍和7额外三连环PBS稀释)被添加到每个实验板进行验证。的平均信号用于所有其他的正常化值在同一个板与Excel软件计算AUC前使用棱镜v.8.4 (GraphPad)。单克隆抗体,欧共体gydF4y2Ba50gydF4y2Ba确定使用的四个参数非线性回归(GraphPad棱镜v.8.4)。gydF4y2Ba
RBD蛋白质的表达gydF4y2Ba
哺乳动物表达载体编码SARS-CoV-2 rbd(基因库MN985325.1;年代蛋白质残基319 - 539)和八个额外突变RBD蛋白质(E484K、Q493R R346S, N493K, N440K, V367F, A475V, S477N和V483A)与一个人类或μ磷酸酶信号肽- 2氨基端以前gydF4y2Ba49gydF4y2Ba。gydF4y2Ba
记者SARS-CoV-2准型病毒gydF4y2Ba
SARS-CoV-2准型粒子生成如前所述gydF4y2Ba1gydF4y2Ba,gydF4y2Ba8gydF4y2Ba。总之,293 t细胞转染pNL4-3ΔEnv-nanoluc和pSARS-CoV-2-SgydF4y2BaΔ19gydF4y2Ba。代RBD-mutant假pSARS-CoV-2-SgydF4y2BaΔ19gydF4y2Ba携带下面的飙升代替其野生型与突变:Q493R, R346S或E484GgydF4y2Ba50gydF4y2Ba。收集粒子在转染后48小时,过滤和储存在−80°C。gydF4y2Ba
准型病毒中和试验gydF4y2Ba
四倍连续稀释等离子体从个人从COVID-19康复的人,或单克隆抗体,孵化了SARS-CoV-2准型病毒1 h在37°C。混合物随后孵化293 tgydF4y2BaACE2gydF4y2Ba细胞48 h后,细胞用PBS和荧光素酶细胞培养细胞溶解消散5×试剂(Promega)。Nanoluc溶解产物荧光素酶活动测量使用Nano-Glo荧光素酶检测系统(Promega) Glomax导航器(Promega)。获得相对发光单元被规范化这些来自细胞感染SARS-CoV-2准型病毒没有等离子体或单克隆抗体。等离子体half-maximal抑制浓度(NTgydF4y2Ba50gydF4y2Ba)或单克隆抗体(ICgydF4y2Ba50gydF4y2Ba)确定使用的四个参数非线性回归(最小二乘回归方法没有权重;约束:顶部= 1,底部= 0)(GraphPad棱镜)。gydF4y2Ba
流式细胞仪数据的高维数据分析gydF4y2Ba
高维viSNE和FlowSOM数据分析和可视化的流式细胞仪数据进行B细胞使用Cytobank平台(gydF4y2Bahttps://cytobank.orggydF4y2Ba)。viSNE分析使用相同抽样的4893个细胞从每个FCS文件,与75年00迭代,30困惑和θ为0.5。以下标记被用来生成viSNE地图:IgA, CD305, TGFb-RII, CD138, CD10, CD272, IgD, CD24, CD21,表征CD95, HLA-DR,免疫球蛋白,CD279, CD38、IgM, CD274, CD27, CD23、CXCR5, CD32, CD86, CD40, CD85j, CD11c和CXCR3。结果viSNE地图被送入FlowSOM聚类算法gydF4y2Ba51gydF4y2Ba。自组织映射生成使用分层集群上的共识gydF4y2BatgydF4y2Ba分布式随机邻居嵌入轴。gydF4y2Ba
热点图可视化gydF4y2Ba
热图显示column-scaledgydF4y2BazgydF4y2Ba分数的平均荧光强度对个人FlowSOM集群根据标记表达式创建pheatmap使用R的函数。gydF4y2Ba
生物素酰化的病毒蛋白用于流式细胞术gydF4y2Ba
纯化,Avi-tagged SARS-CoV-2 RBD是生物素化的使用Biotin-Protein Ligase-BIRA装备根据制造商的指示(活动性)如前所述gydF4y2Ba1gydF4y2Ba。卵清蛋白(σ,A5503-1G)生物素化的使用EZ-Link Sulfo-NHS-LC-Biotinylation装备根据制造商的指示(热科学)。生物素化的卵清蛋白是共轭streptavidin-BV711 (BD生物科学,563262)和RBD streptavidin-PE (BD生物科学,554061)和streptavidin-AF647 (Biolegend, 405237)gydF4y2Ba1gydF4y2Ba。gydF4y2Ba
单细胞排序通过流式细胞术gydF4y2Ba
单细胞通过流式细胞术曾描述排序gydF4y2Ba1gydF4y2Ba。总之,外周血单核细胞B细胞的负选择使用pan-B-cell隔离设备根据制造商的指示(Miltenyi研究,130-101-638)。丰富B细胞孵化在流式细胞仪缓冲区(1×PBS, 2% FCS, 1毫米EDTA)使用以下反抗体(在1:200稀释):anti-CD20-PECy7 (BD生物科学,335793),780年anti-CD3-APC-eFluro(表达载体,47-0037-41),780年anti-CD8-APC-eFluor(表达载体,47-0086-42),780年anti-CD16-APC-eFluor(表达载体,47-0168-41),780年anti-CD14-APC-eFluor(表达载体,47-0149-42),以及僵尸近红外光谱(BioLegend, 423105)和fluorophore-labelled RBD和卵清蛋白(Ova) 30分钟在冰上。单一CD3gydF4y2Ba−gydF4y2BaCD8gydF4y2Ba−gydF4y2BaCD14gydF4y2Ba−gydF4y2BaCD16gydF4y2Ba−gydF4y2BaCD20gydF4y2Ba+gydF4y2Ba卵子gydF4y2Ba−gydF4y2BaRBD-PEgydF4y2Ba+gydF4y2BaRBD-AF647gydF4y2Ba+gydF4y2BaB细胞被分为个别井96 -孔板包含4μl裂解缓冲(0.5×PBS, 10毫米德勤,3000单位/毫升RNasin核糖核酸酶抑制剂(Promega N2615))每口井使用流式细胞仪咏叹调三世和FACSDiva软件(Becton Dickinson)收购和FlowJo进行分析。干冰的排序细胞冷冻,然后储存在−80°C或立即用于随后的RNA逆转录。gydF4y2Ba
抗体测序、克隆和表达gydF4y2Ba
抗体是鉴定和测序如前所述gydF4y2Ba1gydF4y2Ba。总之,从单个细胞RNA是reverse-transcribed(上标三世逆转录酶,表达载体,18080 - 044)和cDNA储存在−20°C或用于后续放大的变量本,IGL和嵌套PCR和桑格IGK基因测序。使用MacVector进行序列分析。从第一个PCR反应扩增子被用作模板序列——和ligation-independent克隆抗体表达向量。重组单克隆抗体和晶圆厂生产和纯化如前所述gydF4y2Ba1gydF4y2Ba。gydF4y2Ba
计算分析抗体序列gydF4y2Ba
抗体序列被削减的基础上质量和注释使用Igblastn v.1.14。IMGT域描述系统。系统地使用Change-O工具箱v.0.4.540执行注释gydF4y2Ba52gydF4y2Ba。重型和轻型链源自相同的细胞配对,和clonotypes被分配的基础上他们的V和J基因内部使用R和Perl脚本(扩展数据图。gydF4y2Ba5gydF4y2Ba)。所有的脚本和数据用于处理抗体序列是公开在GitHub (gydF4y2Bahttps://github.com/stratust/igpipelinegydF4y2Ba)。gydF4y2Ba
人类V基因的频率分布anti-SARS-CoV-2抗体相比,本研究从131284220年以前生成的本和IgL序列gydF4y2Ba53gydF4y2Ba并从cAb-Rep下载gydF4y2Ba54gydF4y2Ba(人类共享BCR clonotypes可用的数据库gydF4y2Bahttps://cab-rep.c2b2.columbia.edu/gydF4y2Ba)。82种不同V基因的基础上,1703年从免疫球蛋白序列分析的三个参与者在这项研究中,我们选择了本和IgL序列从数据库由同一V基因部分编码和统计根据恒定区。频率(扩展数据图所示。gydF4y2Ba5gydF4y2Ba)是相对于源和同形像进行分析。我们使用双面二项测试来检查序列的数量是否属于一个特定IGHV或IGLV曲目的基因是不同的根据相同的频率IGV基因数据库中。调整gydF4y2BaPgydF4y2Ba值计算使用错误发现率修正。在扩展数据无花果。gydF4y2Ba5gydF4y2Ba,gydF4y2Ba6gydF4y2Ba显著差异,标有星号(*gydF4y2BaPgydF4y2Ba< 0.05,* *gydF4y2BaPgydF4y2Ba< 0.01,* * *gydF4y2BaPgydF4y2Ba< 0.001,* * * *gydF4y2BaPgydF4y2Ba< 0.0001)。gydF4y2Ba
核苷酸体细胞hypermutation和CDR3上长度测定使用内部的R和Perl脚本。体细胞hypermutations IGHV和IGLV核苷酸序列一致反对他们的亲密中使用Igblastn和数量的差异被认为是核苷酸突变。计算V的平均变异基因的总和除以所有参与者间核苷酸突变序列的数量用于分析。计算肉汁的疏水性gydF4y2Ba55gydF4y2Ba我们使用了人h . r .疏水性规模基于自由能的转移(千卡每摩尔)gydF4y2Ba56gydF4y2Ba实现的R包肽(gydF4y2Bahttps://CRAN.R-project.org/package=PeptidesgydF4y2Ba)。我们使用532重链CDR3上从这项研究氨基酸序列和22654256本CDR3上序列从公众记忆B细胞受体序列的数据库gydF4y2Ba57gydF4y2Ba。Shapiro-Wilk测试被用来确定肉汁成绩正态分布。肉汁分数从532本CDR3上氨基酸序列从这项研究被用来执行测试和5000年肉汁的公共数据库的序列是随机选择的。的Shapiro-WilkgydF4y2BaPgydF4y2Ba值是6.896×10gydF4y2Ba−3gydF4y2Ba和2.217×10gydF4y2Ba−6gydF4y2Ba序列从这项研究和公共数据库,分别表明数据不是正态分布。因此,我们使用了双尾Wilcoxon非参数检验比较样本,这表明一个疏水性的差异分布(gydF4y2BaPgydF4y2Ba= 5×10gydF4y2Ba−6gydF4y2Ba)(扩展数据图。gydF4y2Ba6小时gydF4y2Ba)。gydF4y2Ba
日志的热图gydF4y2Ba2 gydF4y2Ba改变相对折叠EC的变化gydF4y2Ba50gydF4y2Ba对表示RBD抗体克隆对获得的突变体(图1.3和6.2个月。gydF4y2Ba3 egydF4y2Ba,扩展数据图。gydF4y2Ba7 kgydF4y2Ba创建)R pheatmap包(gydF4y2Bahttps://github.com/raivokolde/pheatmapgydF4y2Ba)使用欧氏距离和Ward.2聚类方法。gydF4y2Ba
活组织检查和免疫荧光gydF4y2Ba
使用内窥镜粘膜活检获得formalin-fixed,石蜡包埋。部分(5μm)被削减,在二甲苯脱蜡,患者在分级酒精和PBS。燥热引起抗原决定基检索在目标执行检索解决方案(DAKO S1699)使用商业压力锅。幻灯片被冷却到室温,洗在PBS和permeabilized 30分钟在PBS 0.1% Triton x - 100。非特异性结合被10%的山羊血清(表达载体,50062 z)在室温下1 h。部分被孵化的主要抗体稀释阻挡一夜之间解决方案在4°C。幻灯片是在PBS洗3次,然后孵化在二级抗体和DAPI(1μg毫升gydF4y2Ba−1gydF4y2Ba在室温下)1 h。部分在PBS清洗三次,然后用Fluoromount-G安装(电子显微镜科学,1798425)。控制包括省略主要抗体(不主995控制)或用初级抗体不反应的抗体相同的同形像(同形像控制)。Eclipse倪尼康显微镜和数码单反相机(尼康,DS-Qi2)被用来组织可视化和图像。gydF4y2Ba
抗体染色部分用于对冠N和N蛋白在兔子长大与SARS-CoV-2 N可交叉反应的蛋白质gydF4y2Ba58gydF4y2Ba(补充表gydF4y2Ba8gydF4y2Ba)。gydF4y2Ba
SARS-CoV-2 PCR从肠活检gydF4y2Ba
确定SARS-CoV-2 RNA存在于胃肠道,我们从粘膜活检获得使用内窥镜使用孤立的RNA Direct-zol miniprep工具包(R2050 Zymo研究)。Reverse-transcribed cDNA放大使用2019 - ncov Ruo工具包(IDT)检测基因组RNA病毒核衣壳。放大的subgenomic核衣壳RNA是使用以下引物和探针:sgLeadSARSCov2_F 5′-CGATCTCTTGTAGATCTGTTCTC-3′gydF4y2Ba28gydF4y2BawtN_P4 wtN_R4 5′-GGTGAACCAAGACGCAGTAT-3′, 5′- / 56-FAM / TAACCAGAA /禅/ TGGAGAACGCAGTGGG / 3 iabkfq / 3′。gydF4y2Ba
使用QuantTect探针定量PCR进行PCR工具包(试剂盒,204345)在以下条件下:95°C 15 s、95°C 15秒和60°C 1分钟使用应用生物系统QuantStudio 6 Flex实时PCR系统。病毒RNA被认为是发现如果gydF4y2BaCgydF4y2BatgydF4y2Ba对病毒的引物和探针组合是< 40岁。正样本井column-purified和N1另外Sanger测序验证了序列的存在。gydF4y2Ba
SARS-CoV-2 RNA的检测探头距离结扎gydF4y2Ba
探针设计与20 - 25 SARS-CoV-2基因组RNA的核苷酸同源性。调查评估通过NCBI BLAST排除掉目标绑定到其他细胞记录。IDT OligoAnalyzer(集成DNA技术)是用于识别探针对相似的热力学性质;熔化温度45 - 60°C, GC含量和低后40 - 55%。3′末端的每一个探头信号放大的目的是用于邻近结扎部分互补序列61 - bp长骨干和部分21个基点插入(补充表gydF4y2Ba8gydF4y2Ba)。单分子荧光原位杂交(smFISH)探针设计与生物素的3′末端互补检测探针(补充表gydF4y2Ba8gydF4y2Ba)。gydF4y2Ba
石蜡包埋在10μm样本分组。部分deparaffinized使用100%二甲苯,5分钟在室温下,重复两次。幻灯片在100%乙醇冲洗,在室温下1分钟,两次,风干。内源性过氧化物酶活性被用0.3%的过氧化氢,治疗样品在室温下10分钟之后,用DEPC-treated水清洗。样本孵化15分钟在抗原95 - 100°C检索解决方案(ACDBio)在DEPC-treated冲洗水和100%乙醇脱水,3分钟在室温和风干。组织部分permeabilized 30分钟在40°C使用RNAscope蛋白酶+解决方案(ACDBio)和DEPC-treated冲洗水。gydF4y2Ba
一夜之间杂交进行在40°C缓冲区根据DEPC-treated水含有2×SSC, 20%甲酰胺(热费希尔科学),2.5% (v / v) polyvinylsulfonic酸,20毫米核糖核苷vanadyl复杂(新英格兰生物学实验室),40 U mlgydF4y2Ba−1gydF4y2BaRNasin (Promega), 0.1% (v / v)渐变20(西格玛奥德里奇),100μg毫升gydF4y2Ba−1gydF4y2Ba大马哈鱼精子DNA(热费希尔科学),100μg毫升gydF4y2Ba−1gydF4y2Ba酵母RNA(热费希尔科学)。DNA探针溶解在DEPC-treated水被添加在最后100海里(集成DNA技术)的浓度。样本洗短暂和孵化缓冲区包含2×SSC, 20%甲酰胺,40 U mlgydF4y2Ba−1gydF4y2BaRNasin 40°C,然后洗了四次(每5分钟)在洗缓冲区,PBS, 0.1% (v / v)渐变20日和4 U mlgydF4y2Ba−1gydF4y2BaRNasin (Promega)。幻灯片被孵化与100 nM插入和骨干寡核苷酸在PBS, 1×SSC, 0.1% (v / v)二层100μg毫升gydF4y2Ba−1gydF4y2Ba大马哈鱼精子DNA(热费希尔科学),100μg毫升gydF4y2Ba−1gydF4y2Ba酵母RNA(热费希尔科学),40 U mlgydF4y2Ba−1gydF4y2BaRNasin在37°C。四个洗后,组织被孵化与0.1 Uμl 37°CgydF4y2Ba−1gydF4y2BaT4 DNA连接酶(新英格兰生物学实验室)在50 mM Tris-HCl MgCl 10毫米gydF4y2Ba2 gydF4y2Ba1毫米ATP, 250毫米德勤,μg毫升gydF4y2Ba−1gydF4y2BaBSA、0.05%渐变20、40 U mlgydF4y2Ba−1gydF4y2BaRNasin,其次是与0.1 Uμl孵化gydF4y2Ba−1gydF4y2Baphi29 DNA聚合酶在50 mM Tris-HCl MgCl 10毫米gydF4y2Ba2 gydF4y2Ba10毫米(NHgydF4y2Ba4gydF4y2Ba)gydF4y2Ba2 gydF4y2Ba所以gydF4y2Ba4gydF4y2Ba250μM核苷酸,1毫米德勤,0.05%渐变20、40 U mlgydF4y2Ba−1gydF4y2BaRNasin pH值7.5 30°C。幻灯片是清洗和使用亲和素、生物素阻断内源性生物素被工具包(向量实验室)根据制造商的指示。滚动循环扩增子被确定使用biotin-labelled DNA探针的浓度5 nM在PBS 37°C, 1×SSC, 0.1%渐变100μg毫升gydF4y2Ba−1gydF4y2Ba大马哈鱼精子DNA、100μg毫升gydF4y2Ba−1gydF4y2Ba酵母RNA。洗后,样本孵化1:10 0稀释streptavidin-HRP在PBS(热费希尔科学),在室温下60分钟洗紧随其后。荧光标签使用Alexa萤石完成647 Tyramide SuperBoostKit(热费希尔科学)根据制造商的指示。赫斯特33342年用于核复染色(热费希尔科学)和样本安装在延长黄金不变色(热费希尔科学)。gydF4y2Ba
由smFISH SARS-CoV-2 RNA检测gydF4y2Ba
一夜之间杂交进行在40°C缓冲区根据DEPC-treated水含有2×SSC, 20%甲酰胺(热费希尔科学),2.5% (v / v) polyvinylsulfonic酸,20毫米核糖核苷vanadyl复杂(新英格兰生物学实验室),40 U mlgydF4y2Ba−1gydF4y2BaRNasin (Promega), 0.1% (v / v)渐变20(西格玛奥德里奇),100μg毫升gydF4y2Ba−1gydF4y2Ba大马哈鱼精子DNA(热费希尔科学),100μg毫升gydF4y2Ba−1gydF4y2Ba酵母RNA(热费希尔科学)。DNA探针溶解在DEPC-treated水被添加在最后一集中10 nM(集成DNA技术)。样本洗短暂和孵化缓冲区包含2×SSC, 20%甲酰胺,40 U mlgydF4y2Ba−1gydF4y2BaRNasin 40°C,然后在洗洗四次缓冲区,PBS, 0.1% (v / v)渐变20日和4 U mlgydF4y2Ba−1gydF4y2BaRNasin (Promega)。样本清洗和使用亲和素、生物素阻断内源性生物素被工具包(向量实验室)根据制造商的指示。幻灯片是孵化与biotin-labelled DNA探针的浓度10 nM在PBS 37°C, 1×SSC, 0.1%渐变100μg毫升gydF4y2Ba−1gydF4y2Ba大马哈鱼精子DNA、100μg毫升gydF4y2Ba−1gydF4y2Ba酵母RNA。洗后,样本孵化1:10 0稀释streptavidin-HRP在PBS(热费希尔科学),在室温下60分钟洗紧随其后。样本的标签使用ImmPACT-DAB衬底,用苏木精复染色QS和嵌入VectaMount AQ安装介质(向量实验室)根据制造商的指示。gydF4y2Ba
数据显示gydF4y2Ba
在2020年Adobe Illustrator扩展数据数据安排。gydF4y2Ba
报告总结gydF4y2Ba
进一步研究信息设计是可用的gydF4y2Ba自然研究报告摘要gydF4y2Ba与本文有关。gydF4y2Ba
数据可用性gydF4y2Ba
数据提供了补充表gydF4y2Ba1gydF4y2Ba- - - - - -gydF4y2Ba8gydF4y2Ba。原始测序数据和计算机脚本与图有关。gydF4y2Ba2 gydF4y2Ba一直在沉积gydF4y2Bahttps://github.com/stratust/igpipelinegydF4y2Ba。这项研究还使用数据从一个森林女神入世与裁判。gydF4y2Ba57gydF4y2Ba(gydF4y2Bahttps://doi.org/10.5061/dryad.35ks2gydF4y2Ba),蛋白质数据银行(加入代码gydF4y2Ba6 vybgydF4y2Ba和gydF4y2Ba6 nb6gydF4y2Ba)和裁判。gydF4y2Ba53gydF4y2Ba。gydF4y2Ba
代码的可用性gydF4y2Ba
计算机代码来处理抗体序列可以在GitHub (gydF4y2Bahttps://github.com/stratust/igpipelinegydF4y2Ba)。gydF4y2Ba
引用gydF4y2Ba
Robbiani, d . f . et al .收敛抗体反应SARS-CoV-2康复的人。gydF4y2Ba自然gydF4y2Ba584年gydF4y2Ba,437 - 442 (2020)。gydF4y2Ba
谢弗,a . et al .抗体效价,组合效应函数,在保护和治疗SARS-CoV-2感染体内。gydF4y2Baj . Exp地中海。gydF4y2Ba。gydF4y2Ba218年gydF4y2Bae20201993 (2021)。gydF4y2Ba
Carfi,。,Bern一个be我,R。&l一个nd我,F. Persistent symptoms in patients after acute COVID-19.j。医学协会。gydF4y2Ba。gydF4y2Ba324年gydF4y2Ba,603 - 605 (2020)。gydF4y2Ba
德尔里奥,C。,Coll我ns, L. F. & Malani, P. Long-term Health Consequences of COVID-19.《美国医学会杂志》gydF4y2Ba324年gydF4y2Ba,1723 - 1724 (2020)。gydF4y2Ba
杨,h . s . et al . SARS-CoV-2抗体特征在急诊室,由两个半定量分析住院和康复的病人。gydF4y2Ba中国。詹。学报gydF4y2Ba509年gydF4y2Ba,117 - 125 (2020)。gydF4y2Ba
罗氏诊断。gydF4y2Ba罗氏诊断gydF4y2BaElecsys Anti-SARS-CoV-2gydF4y2Ba,gydF4y2Bahttps://www.fda.gov/media/137605gydF4y2Ba(2020)。gydF4y2Ba
Muecksch, f . et al .纵向分析COVID19康复者的血清学和中和抗体水平。gydF4y2Baj .感染。说gydF4y2Ba。jiaa659 (2020)。gydF4y2Ba
施密特,f . et al .测量SARS-CoV-2使用准型和嵌合病毒中和抗体活性。gydF4y2Baj . Exp地中海。gydF4y2Ba。gydF4y2Ba217年gydF4y2Bae20201181 (2020)。gydF4y2Ba
Wajnberg, a . et al .健壮的中和抗体SARS-CoV-2感染持续好几个月。gydF4y2Ba科学gydF4y2Ba370年gydF4y2Ba,1227 - 1230 (2020)。gydF4y2Ba
?萧和,j . et al .纵向观察和下降的中和抗体反应后的三个月内SARS-CoV-2感染人类。gydF4y2BaNat。MicrobiolgydF4y2Ba。gydF4y2Ba5gydF4y2Ba,1598 - 1607 (2020)。gydF4y2Ba
这,p . j . m . et al . COVID-19患者有效的中和抗体定义多个目标的脆弱性。gydF4y2Ba科学gydF4y2Ba369年gydF4y2Ba,643 - 650 (2020)。gydF4y2Ba
曹,y等。有效的中和抗体SARS-CoV-2被高通量单细胞测序的恢复期的患者的B细胞。gydF4y2Ba细胞gydF4y2Ba182年gydF4y2Ba,73 - 84 (2020)。gydF4y2Ba
居,b . et al。人类SARS-CoV-2感染引发的中和抗体。gydF4y2Ba自然gydF4y2Ba584年gydF4y2Ba,115 - 119 (2020)。gydF4y2Ba
Kreer, c . et al .纵向隔离的near-germline SARS-CoV-2-neutralizing COVID-19患者的抗体。gydF4y2Ba细胞gydF4y2Ba182年gydF4y2Ba,843 - 854 (2020)。gydF4y2Ba
放弃、大肠等。分析SARS-CoV-2-infected个人揭示了发展的强有力的中和抗体与有限的体细胞突变。gydF4y2Ba免疫力gydF4y2Ba53gydF4y2Ba,98 - 105 (2020)。gydF4y2Ba
元,m . et al。结构共享SARS-CoV-2抗体反应的基础。gydF4y2Ba科学gydF4y2Ba369年gydF4y2Ba,1119 - 1123 (2020)。gydF4y2Ba
Weisblum, y . et al .逃离中和抗体SARS-CoV-2突起蛋白变体。gydF4y2BaeLifegydF4y2Ba9gydF4y2Bae61312 (2020)。gydF4y2Ba
巴恩斯,c . o . et al . SARS-CoV-2中和抗体结构通知治疗策略。gydF4y2Ba自然gydF4y2Ba588年gydF4y2Ba,682 - 687 (2020)。gydF4y2Ba
Tortorici, m . a . et al . Ultrapotent人类抗体预防SARS-CoV-2挑战通过多种机制。gydF4y2Ba科学gydF4y2Ba370年gydF4y2Ba,950 - 957 (2020)。gydF4y2Ba
Baum, a . et al .抗体鸡尾酒SARS-CoV-2突起蛋白阻止快速突变逃避与单个抗体。gydF4y2Ba科学gydF4y2Ba369年gydF4y2Ba,1014 - 1018 (2020)。gydF4y2Ba
李问:et al .突变的影响在病毒性传染性和抗原性SARS-CoV-2飙升。gydF4y2Ba细胞gydF4y2Ba182年gydF4y2Ba,1284 - 1294 (2020)。gydF4y2Ba
拉默斯先生,m . m . et al . SARS-CoV-2有效感染人类肠道肠上皮细胞。gydF4y2Ba科学gydF4y2Ba369年gydF4y2Ba,50 - 54 (2020)。gydF4y2Ba
Livanos, a . e . et al .胃肠道参与变弱COVID-19严重程度和死亡率。预印在gydF4y2Bahttps://doi.org/10.1101/2020.09.07.20187666gydF4y2Ba(2020)。gydF4y2Ba
钱,问:et al。活跃的SARS-CoV-2复制肠的直接证据。gydF4y2Ba中国。感染。说gydF4y2Ba。ciaa925 (2020)。gydF4y2Ba
汉族,y . et al。SARS-CoV-2抑制剂使用肺和结肠瀑样的识别。gydF4y2Ba自然gydF4y2Ba589年gydF4y2Ba,270 - 275 (2021)。gydF4y2Ba
马龙、g . et al .发病率和持久性COVID-19病毒传染的急性患者negativized咽拭子:一个系统的复习。gydF4y2Ba前面。地中海(洛桑)。gydF4y2Ba7gydF4y2Ba562 (2020)。gydF4y2Ba
公园,s . k . et al .粪便样本中检测SARS-CoV-2无症状的患者,在韩国COVID-19温和。gydF4y2Ba中国。杂志。乙醇gydF4y2Ba。gydF4y2Bahttps://doi.org/10.1016/j.cgh.2020.06.005gydF4y2Ba(2020)。gydF4y2Ba
Wolfel, r . et al .病毒学评估住院患者covid - 2019。gydF4y2Ba自然gydF4y2Ba581年gydF4y2Ba,465 - 469 (2020)。gydF4y2Ba
Beaudoin-Bussieres, g . et al .减少体液反应对SARS-CoV-2激增康复的人。gydF4y2BaMBiogydF4y2Ba11gydF4y2Bae02590 - 20398 (2020)。gydF4y2Ba
k·h·d·克劳福德et al .动力学SARS-CoV-2感染后几个月的中和抗体滴度。gydF4y2Baj .感染。说gydF4y2Ba。jiaa618 (2020)。gydF4y2Ba
艾耶a . s . et al .持久性和衰变的人类抗体反应的受体结合域SARS-CoV-2飙升COVID-19患者的蛋白质。gydF4y2Ba科学。ImmunolgydF4y2Ba。gydF4y2Ba5gydF4y2Baeabe0367 - 0313 (2020)。gydF4y2Ba
王,k . et al .纵向动力学的中和抗体反应SARS-CoV-2感染。gydF4y2Ba中国。感染。说gydF4y2Ba。gydF4y2Ba579年gydF4y2Ba,270 - 279 (2020)。gydF4y2Ba
Ripperger, t . j . et al .正交SARS-CoV-2血清学检测使监测低流行率的社区和揭示耐用体液免疫。gydF4y2Ba免疫力gydF4y2Ba53gydF4y2Ba,925 - 933。e4 (2020)。gydF4y2Ba
丹,j . m . et al .免疫记忆SARS-CoV-2评估长达8个月后感染。gydF4y2Ba科学gydF4y2Bahttps://doi.org/10.1126/science.abf4063gydF4y2Ba(2021)。gydF4y2Ba
瑞达,l . b . et al .功能性SARS-CoV-2-specific免疫记忆轻度COVID-19后依然存在。gydF4y2Ba细胞gydF4y2Ba184年gydF4y2Ba,169 - 183 (2021)。gydF4y2Ba
西奇,与免疫球蛋白m . et al . COVID-19生存伙伴应对SARS-CoV-2飙升受体结合域。gydF4y2Baj .中国。投资gydF4y2Ba。gydF4y2Ba130年gydF4y2Ba,6366 - 6378 (2020)。gydF4y2Ba
刘,l . et al。有效的中和抗体上的多个抗原表位SARS-CoV-2飙升。gydF4y2Ba自然gydF4y2Ba584年gydF4y2Ba,450 - 456 (2020)。gydF4y2Ba
Zost, s . j . et al .强有力地人类SARS-CoV-2抗体中和和保护。gydF4y2Ba自然gydF4y2Ba584年gydF4y2Ba,443 - 449 (2020)。gydF4y2Ba
尼尔森,s c a . et al。人类B细胞克隆扩张和收敛SARS-CoV-2抗体反应。gydF4y2Ba宿主细胞活gydF4y2Ba。gydF4y2Ba28gydF4y2Ba,516 - 525 (2020)。gydF4y2Ba
金子:et al .丧失Bcl-6-expressing T卵泡COVID-19辅助细胞和生发中心。gydF4y2Ba细胞gydF4y2Ba183年gydF4y2Ba,143 - 157 (2020)。gydF4y2Ba
Victora、gdp和生理压力,m . c .生发中心。gydF4y2Ba为基础。启ImmunolgydF4y2Ba。gydF4y2Ba30.gydF4y2Ba,429 - 457 (2012)。gydF4y2Ba
索,a . et al .成熟和持久性的anti-SARS-CoV-2记忆B细胞的反应。预印在gydF4y2Bahttps://doi.org/10.1101/2020.11.17.385252gydF4y2Ba(2020)。gydF4y2Ba
马勒,d . a &威尔斯,c . k .评估评级呼吸困难的临床方法。gydF4y2Ba胸部gydF4y2Ba93年gydF4y2Ba,580 - 586 (1988)。gydF4y2Ba
Pujadas、大肠等。SARS-CoV-2病毒载量预测COVID-19死亡率。gydF4y2Ba和出版的《柳叶刀》杂志上。地中海gydF4y2Ba。gydF4y2Ba8gydF4y2Bae70 (2020)。gydF4y2Ba
Chomczynski p &萨基:单步方法的RNA隔离酸胍盐thiocyanate-phenol-chloroform提取。gydF4y2Ba肛交。物化学gydF4y2Ba。gydF4y2Ba162年gydF4y2Ba,156 - 159 (1987)。gydF4y2Ba
DeAngelis, M . M。,Wang, D. G. & Hawkins, T. L. Solid-phase reversible immobilization for the isolation of PCR products.核酸ResgydF4y2Ba。gydF4y2Ba23gydF4y2Ba,4742 - 4743 (1995)。gydF4y2Ba
热那亚,a . et al . T细胞反应的目标与COVID-19 SARS-CoV-2冠状病毒在人类疾病和未暴露的个体。gydF4y2Ba细胞gydF4y2Ba181年gydF4y2Ba,1489 - 1501 (2020)。gydF4y2Ba
Amanat, f . et al .血清学试验检测SARS-CoV-2人类血清转化。gydF4y2Ba地中海Nat。gydF4y2Ba。gydF4y2Ba26gydF4y2Ba,1033 - 1036 (2020)。gydF4y2Ba
巴恩斯,c . o . et al .结构的人类抗体绑定到SARS-CoV-2飙升揭示常见抗体的抗原表位和复发性特征。gydF4y2Ba细胞gydF4y2Ba182年gydF4y2Ba,828 - 842 (2020)。gydF4y2Ba
Weisblum, y . et al .逃离中和抗体SARS-CoV-2突起蛋白变体。gydF4y2BaeLifegydF4y2Ba9gydF4y2Bae61312 (2020)。gydF4y2Ba
Van Gassen, s . et al . FlowSOM:使用自组织映射为可视化和血细胞计数数据的解释。gydF4y2Ba血细胞计数一次gydF4y2Ba87年gydF4y2Ba,636 - 645 (2015)。gydF4y2Ba
古普塔:t . et al . Change-O:工具箱分析大型B细胞免疫球蛋白曲目测序数据。gydF4y2Ba生物信息学gydF4y2Ba31日gydF4y2Ba,3356 - 3358 (2015)。gydF4y2Ba
索托,c . et al .高频的共享人类B细胞受体clonotypes体验。gydF4y2Ba自然gydF4y2Ba566年gydF4y2Ba,398 - 402 (2019)。gydF4y2Ba
郭,Y。,Chen,K., Kwong, P. D., Shapiro, L. & Sheng, Z. cAb-Rep: a database of curated antibody repertoires for exploring antibody diversity and predicting antibody prevalence.前面。ImmunolgydF4y2Ba。gydF4y2Ba10gydF4y2Ba2365 (2019)。gydF4y2Ba
肚子,j . &杜利特尔,r . f .一个简单的方法显示蛋白质的水疗法的字符。gydF4y2Baj·摩尔医学杂志gydF4y2Ba。gydF4y2Ba157年gydF4y2Ba,105 - 132 (1982)。gydF4y2Ba
家伙,h . r .残留的能量和氨基酸侧链分区分布可溶性蛋白质。gydF4y2BaBiophys。JgydF4y2Ba。gydF4y2Ba47gydF4y2Ba,61 - 70 (1985)。gydF4y2Ba
德维特,w . s . et al。一个公共数据库的内存和天真的b细胞受体序列。gydF4y2Ba《公共科学图书馆•综合》gydF4y2Ba11gydF4y2Bae0160853 (2016)。gydF4y2Ba
明镜,m . et al .抑制β干扰素诱导的严重急性呼吸系统综合症冠状病毒显示激活干扰素调节因子的两步模型3。gydF4y2Baj .性研究gydF4y2Ba。gydF4y2Ba79年gydF4y2Ba,2079 - 2086 (2005)。gydF4y2Ba
确认gydF4y2Ba
我们感谢所有参与者学习时间致力于我们的研究;b·科勒和s·施莱辛格,洛克菲勒大学医院临床研究支持办公室和护理人员;大川弗兰克和r·m·b·达内尔SARS-CoV-2唾液PCR检测;c . m .大米和M.C.N.实验室的所有成员有益的讨论;m .扬科维奇实验室支持;和j . Vielmetter蛋白表达中心帮助加州贝克曼研究所的表达式。这项工作是由国家卫生研究院授予P01-AI138398-S1 (M.C.N.和P.J.B.)和2 u19ai111825 (M.C.N.)。加州理工学院的叫法转化研究所和P50 AI150464-13 (P.J.B.),乔治梅森大学快格兰特(D.F.R.和P.J.B.)和欧洲ATAC格兰特EC 101003650 (D.F.R.);R37-AI64003 P.D.B.;R01AI78788 t . Hatziioannou; NCI R01CA234614, NIAID 2R01AI107301, NIDDK R01DK121072 and 1RO3DK117252 to R.E.S., NIH NIDDK R01 DK123749 01S1 to S.M. C.O.B. is supported by the HHMI Hanna Gray and Burroughs Wellcome PDEP fellowships. R.E.S. is an Irma Hirschl Trust Research Award Scholar. M.T. is supported by the Digestive Disease Research Foundation (DDRF). C.G. was supported by the Robert S. Wennett Post-Doctoral Fellowship, in part by the National Center for Advancing Translational Sciences (National Institutes of Health Clinical and Translational Science Award programme, grant UL1 TR001866), and by the Shapiro–Silverberg Fund for the Advancement of Translational Research. P.D.B. and M.C.N. are Howard Hughes Medical Institute Investigators.
作者信息gydF4y2Ba
作者和联系gydF4y2Ba
贡献gydF4y2Ba
C.G.,P。D.B., P.J.B., T. Hatziioannou, S.M. and M.C.N. conceived, designed and analysed the experiments. D.F.R., M. Caskey and C.G. designed clinical protocols. Z.W., J.C.C.L., F.M., S.F., M. Tokuyama, A.C., M.J., D.S.-B., F.S., Y.W., T. Hägglöf, P.M., G.B., C.V., C.O.B., K.G., D.J., J.Y., G.M.-D., Y.B., R.E.S. and Z.Z. carried out experiments. A.G. and M. Cipolla produced antibodies. A.H., D.S.-B., M. Turroja, K.G.M., M. Tankelevich, C.G. and M. Caskey recruited participants and executed clinical protocols. I.S., R.P, J.D. and C.U.-O. processed clinical samples. T.Y.O. and V.R. performed bioinformatic analysis. C.G, P.D.B., P.J.B., T. Hatziioannou, S.M. and M.C.N. wrote the manuscript with input from all co-authors.
相应的作者gydF4y2Ba
道德声明gydF4y2Ba
相互竞争的利益gydF4y2Ba
洛克菲勒大学提起临时专利申请与本工作有关的D.F.R.和M.C.N.发明家(美国专利63/021,387)。R.E.S. Miromatrix公司的科学顾问委员会,是一个顾问和发言人Alnylam Inc .克里担任顾问武田制药形态学和葛兰素史密斯克莱恩。Z.Z.收到种子从等医疗仪器和赞助旅行。gydF4y2Ba
额外的信息gydF4y2Ba
同行审查的信息gydF4y2Ba自然gydF4y2Ba谢谢迈克尔·劳厄斯坦利·帕尔曼和其他匿名的,审稿人(s)为他们的贡献的同行评审工作。gydF4y2Ba
出版商的注意gydF4y2Ba施普林格自然保持中立在发表关于司法主权地图和所属机构。gydF4y2Ba
扩展数据数据和表gydF4y2Ba
扩展数据图1等离子体的临床相关抗体滴定度。gydF4y2Ba
一个gydF4y2Ba,标准化的AUC anti-RBD免疫球蛋白滴定度在1.3个月的参与者(gydF4y2BangydF4y2Ba= 38)或没有(gydF4y2BangydF4y2Ba= 49)持续的急性症状。gydF4y2BabgydF4y2Ba,标准化的AUC anti-RBD免疫球蛋白滴定度在6.2个月的参与者(gydF4y2BangydF4y2Ba= 38)或没有(gydF4y2BangydF4y2Ba= 49)持续的急性症状。gydF4y2BacgydF4y2Ba,标准化的AUC anti-RBD IgM滴定度在1.3个月的参与者(gydF4y2BangydF4y2Ba= 38)或没有(gydF4y2BangydF4y2Ba= 49)持续的急性症状。gydF4y2BadgydF4y2Ba,标准化的AUC anti-RBD IgM滴定度在6.2个月的参与者(gydF4y2BangydF4y2Ba= 38)或没有(gydF4y2BangydF4y2Ba= 49)持续的急性症状。gydF4y2BaegydF4y2Ba,标准化的AUC anti-RBD IgA滴定度在1.3个月的参与者(gydF4y2BangydF4y2Ba= 38)或没有(gydF4y2BangydF4y2Ba= 49)持续的急性症状。gydF4y2BafgydF4y2Ba,标准化的AUC anti-RBD IgA滴定度在6.2个月的参与者(gydF4y2BangydF4y2Ba= 38)或没有(gydF4y2BangydF4y2Ba= 49)持续的急性症状。gydF4y2BaggydF4y2Ba,标准化的AUC anti-N免疫球蛋白滴定度在1.3个月的参与者(gydF4y2BangydF4y2Ba= 38)或没有(gydF4y2BangydF4y2Ba= 49)持续的急性症状。gydF4y2BahgydF4y2Ba,标准化的AUC anti-N免疫球蛋白滴定度在6.2个月的参与者(gydF4y2BangydF4y2Ba= 38)或没有(gydF4y2BangydF4y2Ba= 49)持续的急性症状。gydF4y2Ba我gydF4y2Ba索引值(IV) anti-RBD免疫球蛋白滴定度在1.3个月的参与者(gydF4y2BangydF4y2Ba= 38)或没有(gydF4y2BangydF4y2Ba= 49)持续的急性症状。gydF4y2BajgydF4y2Ba的索引值anti-RBD免疫球蛋白滴定度在6.2个月的参与者(gydF4y2BangydF4y2Ba= 38)或没有(gydF4y2BangydF4y2Ba= 49)持续的急性症状。gydF4y2BakgydF4y2Ba,截止指数(COI)值anti-N总检察长的滴定度在1.3个月的参与者(gydF4y2BangydF4y2Ba= 38)或没有(gydF4y2BangydF4y2Ba= 49)持续的急性症状。gydF4y2BalgydF4y2BaCOI价值观anti-N总检察长的滴定度在6.2个月的参与者(gydF4y2BangydF4y2Ba= 38)或没有(gydF4y2BangydF4y2Ba= 49)持续的急性症状。gydF4y2Ba米gydF4y2Ba,元gydF4y2Ba50gydF4y2Ba值在1.3个月的参与者(gydF4y2BangydF4y2Ba= 38)或没有(gydF4y2BangydF4y2Ba= 49)持续的急性症状。gydF4y2BangydF4y2Ba,元gydF4y2Ba50gydF4y2Ba值在6.2个月的参与者(gydF4y2BangydF4y2Ba= 38)或没有(gydF4y2BangydF4y2Ba= 49)持续的急性症状。gydF4y2BaogydF4y2Ba多年来,年龄的参与者(gydF4y2BangydF4y2Ba= 38)或没有(gydF4y2BangydF4y2Ba= 49)持续的急性症状。gydF4y2BapgydF4y2Ba急性感染的严重程度,评估由世卫的序数临床进展/改进规模参与者(gydF4y2BangydF4y2Ba= 38)或没有(gydF4y2BangydF4y2Ba= 49)持续的急性症状。gydF4y2Ba问gydF4y2Ba的参与者,在急性感染症状的持续时间(gydF4y2BangydF4y2Ba= 38)或没有(gydF4y2BangydF4y2Ba= 49)持续的急性症状。单杠表示中位数的值。使用双尾Mann-Whitney确定统计学意义gydF4y2BaUgydF4y2Ba测试。gydF4y2Ba
扩展数据图2测量等离子体抗体的相关性。gydF4y2Ba
一个gydF4y2Ba,标准化的AUC免疫球蛋白anti-RBD密谋反对塔免疫球蛋白anti-RBD索引值在1.3个月。gydF4y2BabgydF4y2Ba,标准化的AUC免疫球蛋白anti-RBD密谋反对塔免疫球蛋白anti-RBD索引值在6.2个月。gydF4y2BacgydF4y2Ba,标准化的AUC IgM anti-RBD密谋反对塔IgM anti-RBD索引值在1.3个月。gydF4y2BadgydF4y2Ba,标准化的AUC IgM anti-RBD密谋反对塔IgM anti-RBD索引值在6.2个月。gydF4y2BaegydF4y2Ba,规范化的AUC免疫球蛋白anti-N策划对罗氏COI值anti-N总Ig滴定度在1.3个月。gydF4y2BafgydF4y2Ba,规范化的AUC免疫球蛋白anti-N策划对罗氏COI值anti-N总Ig滴定度在6.2个月。gydF4y2BaggydF4y2Ba,Anti-N总Ig COI值80个人在初始1.3 - 6.2月后续访问。gydF4y2BahgydF4y2Ba相对变化anti-N总搞笑水平在1.3和6.2个月策划反对anti-N总搞笑水平1.3个月。gydF4y2Ba我gydF4y2Ba在6.2个月,规范化的AUC IgM anti-RBD策划与IgM anti-RBD 1.3个月。gydF4y2BajgydF4y2Ba6.2个月的免疫球蛋白g anti-RBD规范化AUC策划与免疫球蛋白g anti-RBD 1.3个月。gydF4y2BakgydF4y2Ba在6.2个月,规范化的AUC IgA anti-RBD策划对IgA anti-RBD在1.3个月。gydF4y2BalgydF4y2Ba6.2个月的免疫球蛋白g anti-N规范化AUC策划与免疫球蛋白g anti-N 1.3个月。gydF4y2Ba米gydF4y2Ba,anti-N COI值总Ig滴定度在6.2个月谋害anti-N总Ig滴定度在1.3个月。gydF4y2BangydF4y2Ba,Anti-RBD IgM滴定度在1.3个月谋害anti-N免疫球蛋白滴定度在1.3个月。gydF4y2BaogydF4y2Ba,Anti-RBD免疫球蛋白滴定度在1.3个月谋害anti-N免疫球蛋白滴定度在1.3个月。gydF4y2BapgydF4y2Ba,Anti-RBD IgA滴定度在1.3个月谋害anti-N免疫球蛋白滴定度在1.3个月。gydF4y2Ba问gydF4y2Ba,元gydF4y2Ba50gydF4y2Ba值在1.3个月谋害anti-RBD免疫球蛋白滴定度在1.3个月。gydF4y2BargydF4y2Ba,元gydF4y2Ba50gydF4y2Ba值在6.2个月谋害anti-RBD免疫球蛋白滴定度在6.2个月。的gydF4y2BargydF4y2Ba和gydF4y2BaPgydF4y2Ba值是由双尾枪兵的相关性。gydF4y2Ba
图3扩展数据持久纵向的B细胞表型的变化格局个人从COVID-19中恢复过来。gydF4y2Ba
一个gydF4y2Ba汇集全球viSNE投影B细胞对所有参与者池背景轮廓图所示,连接覆盖预测的控制,和康复的参与者在1.3和6.2个月。gydF4y2BabgydF4y2BaviSNE投影,汇集所有参与者的B细胞B细胞集群被FlowSOM集群。Column-scaledgydF4y2BazgydF4y2Ba分数的平均荧光强度(MFI)的集群和标记。gydF4y2BacgydF4y2Ba从每组、B细胞频率FlowSOM集群表示。每个圆圈代表一个单独的控制个人(gydF4y2BangydF4y2Ba= 20)(灰色),恢复期的参与者在1.3月感染后(gydF4y2BangydF4y2Ba= 41)(红色)或康复的参与者在感染后6.2个月(gydF4y2BangydF4y2Ba= 41)(绿色)。单杠表示平均值。由双尾搭配意义gydF4y2BatgydF4y2Ba之间的比较以及在个人和双尾未配对的时间点gydF4y2BatgydF4y2Ba以及比较控制和康复的人。gydF4y2BadgydF4y2Ba、个人viSNE表示蛋白质的预测表达式。gydF4y2Ba
扩展数据图4流式细胞术。gydF4y2Ba
一个gydF4y2Ba控制策略用于细胞分选。闸门在CD20的汗衫gydF4y2Ba+gydF4y2Ba和CD3gydF4y2Ba−gydF4y2BaCD8gydF4y2Ba−gydF4y2BaCD16gydF4y2Ba−gydF4y2Ba卵子gydF4y2Ba−gydF4y2Ba。细胞被RBD-PE排序gydF4y2Ba+gydF4y2Ba和RBD-AF647gydF4y2Ba+gydF4y2Ba。gydF4y2BabgydF4y2Ba,流式细胞仪显示的百分比RBD-double-positive记忆B细胞从1.3月或6月感染后在21个随机选择的参与者。gydF4y2Ba
扩展数据图5 V的人类基因频率分布。gydF4y2Ba
一个gydF4y2Ba,双面二项测试是用来比较的人类V基因频率分布anti-SARS-CoV-2抗体从捐赠者在1.3个月和6.2个月gydF4y2Ba1gydF4y2Ba。*gydF4y2BaPgydF4y2Ba< 0.05,* *gydF4y2BaPgydF4y2Ba< 0.01 * * *gydF4y2BaPgydF4y2Ba< 0.001,* * * *gydF4y2BaPgydF4y2Ba< 0.0001。gydF4y2BabgydF4y2Ba,双面二项测试是用来比较的人类V基因频率分布anti-SARS-CoV-2抗体从这项研究从参考序列。gydF4y2Ba53gydF4y2Ba。*gydF4y2BaPgydF4y2Ba< 0.05,* *gydF4y2BaPgydF4y2Ba< 0.01,* * *gydF4y2BaPgydF4y2Ba< 0.001,* * * *gydF4y2BaPgydF4y2Ba< 0.0001。gydF4y2BacgydF4y2Ba与克隆、序列从六个人关系描绘成圆环阴谋在无花果。gydF4y2Ba二维gydF4y2Ba。互连线表示抗体之间的关系分享V和J在本和IGL gene-segment序列。紫色、绿色和灰色线连接相关的克隆,克隆和单打,分别和单打。gydF4y2BadgydF4y2Ba为每个参与者,免疫球蛋白重链序列的数量(黑)分析6.2 1.3(左)或月感染后(右)。内部圈子中的数字表示排序为每个单独的细胞的数量高于圆表示。gydF4y2BaegydF4y2Ba一样,gydF4y2BadgydF4y2Ba但是显示所有六个参与者的数据。gydF4y2BafgydF4y2Ba,比较IgG-positive B细胞的百分比从6个人月1.3或6.2月感染后。横条指示的意思。统计学意义是决定使用双尾gydF4y2BatgydF4y2Ba以及。gydF4y2Ba
扩展数据图6分析抗体克隆体细胞hypermutation坚持,CDR3上长度和疏水性。gydF4y2Ba
一个gydF4y2Ba- - - - - -gydF4y2BafgydF4y2Ba、体细胞数核苷酸突变IGVH和IGVL持久化克隆发现月- 1.3月- 6.2时间点在六个参与者:COV21 (gydF4y2Ba一个gydF4y2Ba),COV47 (gydF4y2BabgydF4y2Ba),COV57 (gydF4y2BacgydF4y2Ba),COV72 (gydF4y2BadgydF4y2Ba),COV96 (gydF4y2BaegydF4y2Ba)和COV107 (gydF4y2BafgydF4y2Ba)。每个克隆扩张的VH和重要基因使用上述图表,或表示“汗衫”如果持久化序列是孤立的一次在两个时间点。连接线的体细胞hypermutation克隆对被表示为一个重组单克隆抗体。gydF4y2BaggydF4y2Ba为每个单独的,氨基酸的长度CDR3上IGVH IGVL显示。横条指示的意思。抗体序列的数量(IGVH和IGVL)评估每个参与者gydF4y2BangydF4y2Ba= 90 (COV21),gydF4y2BangydF4y2Ba= 78 (COV47),gydF4y2BangydF4y2Ba= 53 (COV57),gydF4y2BangydF4y2Ba= 87 (COV72),gydF4y2BangydF4y2Ba= 104 (COV96),gydF4y2BangydF4y2Ba= 120 (COV107)。对,所有抗体结合(gydF4y2BangydF4y2BaIGVH和IGVL = 532)。使用双尾Mann-Whitney确定统计学意义gydF4y2BaUgydF4y2Ba测试和单杠表明中值值。NS,不显著,*gydF4y2BaPgydF4y2Ba< 0.05,* *gydF4y2BaPgydF4y2Ba< 0.01,* * *gydF4y2BaPgydF4y2Ba< 0.001,* * * *gydF4y2BaPgydF4y2Ba< 0.0001。gydF4y2BahgydF4y2Ba、分布的疏水性肉汁分数在532年本CDR3上抗体序列相比,本研究从公共数据库:统计分析提供了方法。盒子限制是低级和高级质量,中心行表示中位数,胡须是1.5×四分位范围和点代表离群值。统计学意义是决定使用双尾Wilcoxon配对符号秩检验。NS,不显著,*gydF4y2BaPgydF4y2Ba< 0.05,* *gydF4y2BaPgydF4y2Ba< 0.01,* * *gydF4y2BaPgydF4y2Ba< 0.001,* * * *gydF4y2BaPgydF4y2Ba< 0.0001)。gydF4y2Ba
扩展数据图7 ELISA野生型和突变体RBD单克隆抗体。gydF4y2Ba
一个gydF4y2Ba,电子商务gydF4y2Ba50gydF4y2Ba值绑定到野生型RBD的共享汗衫和共享克隆获得的单克隆抗体在初始1.3 - 6.2月复诊,除以参与者(gydF4y2BangydF4y2Ba= 6 (COV21),gydF4y2BangydF4y2Ba= 13 (COV47),gydF4y2BangydF4y2Ba= 3 (COV57),gydF4y2BangydF4y2Ba= 6 (COV72),gydF4y2BangydF4y2Ba= 15 (COV96),gydF4y2BangydF4y2Ba= 9 (COV107))。线连接共享汗衫或克隆。单克隆抗体与EC的改善gydF4y2Ba50gydF4y2Ba在6.2月的随访中突出显示绿色;剩余的单克隆抗体所示黑色。统计学意义是决定使用双尾Wilcoxon配对符号秩检验。gydF4y2BabgydF4y2Ba- - - - - -gydF4y2BajgydF4y2Ba、图形显示不同的抗体ELISA绑定曲线获得1.3个月(虚线)和他们的克隆获得的亲戚后6.2个月(实线)绑定到野生型,R346S, E484K, Q493R, N439K, N440K, A475V, S477N, V483A V367F rbd(颜色表示)。抗体对的标识符表示的顶部面板(1.3个月和6.2个月)。gydF4y2BakgydF4y2Ba、热地图显示日志gydF4y2Ba2 gydF4y2Ba改变相对折叠EC的变化gydF4y2Ba50gydF4y2Ba对指定的RBD突变体52抗体克隆对获得1.3(黑)和6.2个月(红色)。每个抗体的克隆起源和参与者对上面的指示。gydF4y2Ba
扩展数据图8中和的野生型和突变rbd, C51校准和约束力的投影。gydF4y2Ba
一个gydF4y2Ba,集成电路gydF4y2Ba50gydF4y2Ba值共享汗衫和共享克隆获得的单克隆抗体在最初的1.3和6.2个月的随访中,除以参与者(gydF4y2BangydF4y2Ba= 6 (COV21),gydF4y2BangydF4y2Ba= 13 (COV47),gydF4y2BangydF4y2Ba= 3 (COV57),gydF4y2BangydF4y2Ba= 6 (COV72),gydF4y2BangydF4y2Ba= 15 (COV96),gydF4y2BangydF4y2Ba= 9 (COV107))。线连接共享汗衫或克隆。单克隆抗体检测不到ICgydF4y2Ba50gydF4y2Ba10点在1.3个月策划μg毫升gydF4y2Ba−1gydF4y2Ba用红色突出显示;monoclobal抗体与改善集成电路gydF4y2Ba50gydF4y2Ba在6.2月的随访中突出显示绿色;剩余的单克隆抗体所示黑色。统计学意义是决定使用双尾Wilcoxon配对符号秩检验。gydF4y2BabgydF4y2Ba- - - - - -gydF4y2BafgydF4y2Ba细胞溶解产物,归一化相对发光值293 tgydF4y2BaACE2gydF4y2Ba细胞感染后48 h SARS-CoV-2 pseudovirus包含野生型RBD或RBD突变体(野生型、Q493R E484G和R346S RBD所示黑色,红色,绿色和蓝色,分别)的单克隆抗体浓度增加了在1.3月初始访问(1.3米,虚线)和他们共享克隆或汗衫在6.2月的随访中(6.2,连续线)。抗体标识符表示。gydF4y2BaggydF4y2Ba,VH和重要的氨基酸序列比对C144和导数抗体C051 C052, C053 C054。Germline-encoded残留在绿色突出显示。残留的邻近RBD-binding C144抗体结合部位以红色突出显示。gydF4y2BahgydF4y2Ba- - - - - -gydF4y2BajgydF4y2Ba两个相邻的表面表示“下”RBD (RBD)gydF4y2Ba一个gydF4y2Ba和RBDgydF4y2BaBgydF4y2Ba)飙升的C144抗原决定基三聚物中强调的rbd青色和立场的氨基酸突变积累C052 (gydF4y2BahgydF4y2Ba),C053 (gydF4y2Ba我gydF4y2Ba)和C054 (gydF4y2BajgydF4y2Ba),而父抗体C144,强调把侧链在Cαatom表示。C052, C053 C054与两个rbd的交互建模的基础上低温电子显微镜C144工厂一定会飙升三聚物的结构gydF4y2Ba18gydF4y2Ba。gydF4y2Ba
扩展数据图9 SARS-CoV-2人类肠上皮细胞抗原在胃肠道COVID-19诊断三个月后,没有可检测SARS-CoV-2抗原和pre-COVID-19控制个人。gydF4y2Ba
一个gydF4y2Ba- - - - - -gydF4y2BajgydF4y2Ba人类胃肠组织显示,免疫荧光图像。DAPI染色是EPCAM(红色)(蓝色)和SARS-CoV-2核衣壳(绿色)样本来自肠活检在胃肠道。gydF4y2Ba一个gydF4y2Ba- - - - - -gydF4y2BahgydF4y2Ba活检,从参与者cgi - 088(补充表gydF4y2Ba7gydF4y2Ba)92 d后COVID-19症状。gydF4y2Ba我gydF4y2Ba,gydF4y2BajgydF4y2Ba,活检27个月COVID-19发作前的症状从参与者cgi - 088。箭头表示可检测到SARS-CoV-2肠上皮细胞抗原。每个组织同形像和no-primary控件显示在右两列。酒吧,规模100μm。gydF4y2BakgydF4y2Ba,gydF4y2BalgydF4y2Ba免疫荧光图像的活检样本胃肠道(回肠(gydF4y2BakgydF4y2Ba)和十二指肠(gydF4y2BalgydF4y2Ba)从十pre-COVID-19控制个人获得2018年1月至2019年10月。染色是EPCAM(红色),DAPI(蓝色)和SARS-CoV-2核衣壳(绿色)。酒吧,规模100μm。所有的实验都重复至少两次独立得到了类似的结果。gydF4y2Ba
扩展数据图10 SARS-CoV-2抗原和RNA检测在不同肠段的几个人从COVID-19康复的。gydF4y2Ba
一个gydF4y2Ba、免疫荧光图像的活检样本在胃肠道不同的个体。染色是EPCAM(红色),DAPI(蓝色)和SARS-CoV-2核衣壳(绿色)。样本来自肠活检从四个参与者(cgi - 089, cgi - 092, cgi - 100和cgi - 106)采取COVID-19感染后至少三个月。箭头表示可检测到SARS-CoV-2肠上皮细胞抗原。酒吧,规模100μm。至少两次独立实验重复类似的结果。gydF4y2BabgydF4y2Ba通过免疫荧光、量化SARS-CoV-2-positive细胞。细胞的数量的N蛋白染色阳性SARS-CoV-2每毫米gydF4y2Ba2 gydF4y2Ba肠道上皮细胞。图表显示表示个人的十二指肠活检样本(左)和末端回肠(右)。黑点代表可用活检标本的数量为每个单独的从各自的肠段(cgi - 088,gydF4y2BangydF4y2Ba= 4十二指肠溃疡和gydF4y2BangydF4y2Ba= 2回肠;cgi - 089,gydF4y2BangydF4y2Ba= 2十二指肠溃疡和gydF4y2BangydF4y2Ba= 2回肠;cgi - 092,gydF4y2BangydF4y2Ba= 6十二指肠溃疡和gydF4y2BangydF4y2Ba= 3回肠;cgi - 106,gydF4y2BangydF4y2Ba= 4;cgi - 100,gydF4y2BangydF4y2Ba= 4)。盒子代表值和胡须95%置信区间中值。gydF4y2BacgydF4y2Ba,gydF4y2BadgydF4y2Ba,SARS-CoV-2病毒RNA在肠活检可视化的参与者cgi - 089 (gydF4y2BacgydF4y2Ba)和cgi - 088 (gydF4y2BadgydF4y2Ba用原位杂交)。SARS-CoV-2基因组RNA(黑色)和苏木精和伊红染色smFISH-immunohistochemistry技术在十二指肠(左)或回肠末端(右)。箭头表示肠上皮细胞与探测SARS-CoV-2 RNA。gydF4y2BaegydF4y2BaPre-COVID-19控制人没有检测到SARS-CoV-2病毒RNA在十二指肠(左)或回肠(右)活检。酒吧,规模100μm。实验重复三次独立得到了类似的结果。gydF4y2Ba
补充信息gydF4y2Ba
补充表1gydF4y2Ba
队列研究总结。gydF4y2Ba
补充表2gydF4y2Ba
个别参与者的特征。gydF4y2Ba
补充表3gydF4y2Ba
抗体序列从1.3和6.2月的时间点。gydF4y2Ba
补充表4gydF4y2Ba
序列的克隆单克隆抗体。gydF4y2Ba
补充表5gydF4y2Ba
一半最大有效浓度(EC50s)的单克隆抗体。gydF4y2Ba
补充表6gydF4y2Ba
单克隆抗体的抑制浓度。gydF4y2Ba
补充表7gydF4y2Ba
胃肠道活检组特征。gydF4y2Ba
补充表8gydF4y2Ba
抗体用于免疫荧光和邻近结扎/ smFISH探针序列。gydF4y2Ba
权利和权限gydF4y2Ba
关于这篇文章gydF4y2Ba

引用这篇文章gydF4y2Ba
Gaebler C。王,Z。,lorenz我,J。C。C。gydF4y2Baet al。gydF4y2Ba抗体免疫SARS-CoV-2的进化。gydF4y2Ba自然gydF4y2Ba591年gydF4y2Ba,639 - 644 (2021)。https://doi.org/10.1038/s41586 - 021 - 03207 - wgydF4y2Ba
收到了gydF4y2Ba:gydF4y2Ba
接受gydF4y2Ba:gydF4y2Ba
发表gydF4y2Ba:gydF4y2Ba
发行日期gydF4y2Ba:gydF4y2Ba
DOIgydF4y2Ba:gydF4y2Bahttps://doi.org/10.1038/s41586 - 021 - 03207 - wgydF4y2Ba
本文引用的gydF4y2Ba
体液免疫SARS-CoV-2持久控制及其变体gydF4y2Ba
炎症和再生gydF4y2Ba(2023)gydF4y2Ba
癌症和COVID-19:解体的免疫相互作用的有前途的治疗对严重SARS-CoV-2癌症患者gydF4y2Ba
血液学和肿瘤学杂志》上gydF4y2Ba(2023)gydF4y2Ba
非典CoV-2发现新生儿的大便里远离母亲怀孕期间COVID-19gydF4y2Ba
儿科研究gydF4y2Ba(2023)gydF4y2Ba
长COVID:主要发现,机制和建议gydF4y2Ba
自然评论微生物学gydF4y2Ba(2023)gydF4y2Ba
灭活的疫苗诱导的抗体可以广泛中和SARS-CoV-2循环变量gydF4y2Ba
自然通讯gydF4y2Ba(2023)gydF4y2Ba
评论gydF4y2Ba
通过提交评论你同意遵守我们的gydF4y2Ba条款gydF4y2Ba和gydF4y2Ba社区指导原则gydF4y2Ba。如果你发现一些滥用或不符合我们的条件或准则请国旗是不合适的。gydF4y2Ba
