- 研究gydF4y2Ba
- 开放获取gydF4y2Ba
- 发表:gydF4y2Ba
Akkermansia muciniphilagydF4y2Ba改善老年结肠粘液厚度下降,减弱免疫激活加速老化gydF4y2BaErcc1gydF4y2Ba−/Δ7gydF4y2Ba老鼠gydF4y2Ba
免疫和老化gydF4y2Ba体积gydF4y2Ba16gydF4y2Ba文章编号:gydF4y2Ba6gydF4y2Ba(gydF4y2Ba2019年gydF4y2Ba)gydF4y2Ba
文摘gydF4y2Ba
背景gydF4y2Ba
的使用gydF4y2BaAkkermansia muciniphilagydF4y2Ba作为潜在的治疗干预是收到越来越多的关注。健康益处归功于这种细菌包括改善代谢紊乱和发挥抗炎作用。丰富的gydF4y2Ba答:muciniphilagydF4y2Ba与一个健康的肠道在早期和中期以后的生活。然而,的影响gydF4y2Ba答:muciniphilagydF4y2Ba对肠道健康的下降在老化过程中还没有调查。我们补充加速老化gydF4y2BaErcc1gydF4y2Ba−/Δ7gydF4y2Ba老鼠gydF4y2Ba答:muciniphilagydF4y2Ba10周,进行组织学、转录和肠道健康的免疫方面。gydF4y2Ba
结果gydF4y2Ba
结肠黏液层的厚度增加长期后大约三倍gydF4y2Ba答:muciniphilagydF4y2Ba补充,甚至明显厚相比,老鼠补充gydF4y2Ba乳杆菌gydF4y2BaWCFS1。结肠基因表达谱指向表达减少炎症和免疫功能相关的基因和通路,并建议减少结肠中B细胞的存在。总在脾脏和肠系膜淋巴结B细胞频率没有改变gydF4y2Ba答:muciniphilagydF4y2Ba补充。成熟和不成熟的B细胞在骨髓频率增加,而B细胞前体未受影响。这些发现表明,B细胞迁移而不是生产的影响gydF4y2Ba答:muciniphilagydF4y2Ba补充。基因表达谱在回肠指向降低代谢和免疫相关流程和抗菌肽生产后gydF4y2Ba答:muciniphilagydF4y2Ba补充。除此之外,gydF4y2Ba答:muciniphilagydF4y2Ba激活CD80的频率下降gydF4y2Ba+gydF4y2BaCD273gydF4y2Ba−gydF4y2Ba派尔集合淋巴结补丁的B细胞。此外,腹膜常驻巨噬细胞数量的增加和减少Ly6CgydF4y2BaintgydF4y2Ba单核细胞在脾脏和肠系膜淋巴结频率增加潜在的抗炎作用的证据gydF4y2Ba答:muciniphilagydF4y2Ba。gydF4y2Ba
结论gydF4y2Ba
总之,我们表明,补充gydF4y2Ba答:muciniphilagydF4y2Ba预防结肠黏液层的厚度与年龄相关的下降和减毒炎症和免疫相关流程在老的年龄。这项研究表明gydF4y2Ba答:muciniphilagydF4y2Ba补充有助于促进健康老龄化。gydF4y2Ba
背景gydF4y2Ba
重合的老年人口增加观察到如今,老年人经常不可避免的下降的整体健康正成为一个令人担忧的问题。衰老过程是伴随着慢性低度炎症状态,称为“inflamm-aging”,这是一个强大的风险因素很多老年性疾病(gydF4y2Ba1gydF4y2Ba,gydF4y2Ba2gydF4y2Ba,gydF4y2Ba3gydF4y2Ba,gydF4y2Ba4gydF4y2Ba]。一个器官的衰老过程的影响是肠道和gut-related障碍的发生在老年人人口相当大的gydF4y2Ba5gydF4y2Ba]。gydF4y2Ba
为主要居民的肠道,肠道微生物群的维护起着关键作用的整体健康。细菌能够降解复杂的碳水化合物,从而将这些底物转化为有益健康的代谢物,如短链脂肪酸(SCFAs) (gydF4y2Ba6gydF4y2Ba,gydF4y2Ba7gydF4y2Ba]。此外,肠道微生物群广泛交流与宿主免疫系统的调节免疫反应(gydF4y2Ba8gydF4y2Ba]。在老化过程中,肠道微生物群的成分发生变化,如减少多样性,促进健康的细菌和减少潜在pathobionts的增加。这种干扰微生物群的平衡组成被认为增加肠道屏障功能受损的风险和肠道炎症gydF4y2Ba9gydF4y2Ba]。在老鼠中,将微生物群从老鼠到年轻岁无菌的接受者老鼠提升肠道炎症,增加细菌泄漏组件到血液和刺激系统性免疫激活(gydF4y2Ba10gydF4y2Ba]。gydF4y2Ba
对肠道健康的一个重要因素是黏液层覆盖在肠道上皮细胞层和作为物理保护细菌渗透和有害化合物进入粘膜组织(gydF4y2Ba11gydF4y2Ba]。肠道粘液的建立主要是粘蛋白2 (Muc2)的蛋白质,这是大gel-forming由杯状细胞分泌的蛋白质位于肠道粘膜。这些蛋白质形成的基石是呈网状结构和黏液层。黏液层的重要性强调使用Muc2基因敲除小鼠的研究中,没有一个结肠黏液层覆盖肠道上皮细胞层(gydF4y2Ba12gydF4y2Ba,gydF4y2Ba13gydF4y2Ba]。这些老鼠遭受减少肠道屏障功能,增加炎症状态(gydF4y2Ba14gydF4y2Ba和有结肠炎的迹象gydF4y2Ba12gydF4y2Ba]。黏液层的防护功能,它还作为能量来源的细菌。gydF4y2BaAkkermansia muciniphilagydF4y2Ba是一种细菌物种,能够降低粘液。这种细菌是非常丰富(~ 3%)在人类结肠健康gydF4y2Ba15gydF4y2Ba]。在粘液退化,gydF4y2Ba答:muciniphilagydF4y2Ba产生一些刺激的化合物,如SCFAs和菌毛gydF4y2Ba16gydF4y2Ba,gydF4y2Ba17gydF4y2Ba]。外膜蛋白质pili-like Amuc_1100被认为是参与有益的特性gydF4y2Ba答:muciniphilagydF4y2Ba对健康(gydF4y2Ba18gydF4y2Ba,gydF4y2Ba19gydF4y2Ba]。gydF4y2Ba
最近的研究表明的有利影响gydF4y2Ba答:muciniphilagydF4y2Ba并不局限于肠道,但扩展到整体健康。丰富的gydF4y2Ba答:muciniphilagydF4y2Ba减少人们患肥胖、2型糖尿病、炎症性肠病,在别人(gydF4y2Ba20.gydF4y2Ba]。此外,补充和gydF4y2Ba答:muciniphilagydF4y2Ba在老鼠身上导致一种改进的代谢状态,并减少食源性肥胖(ClinicalTrials.gov标识符:NCT02637115) [gydF4y2Ba21gydF4y2Ba,gydF4y2Ba22gydF4y2Ba,gydF4y2Ba23gydF4y2Ba]。gydF4y2Ba
我们之前和其他人显示丰富的gydF4y2BaAkkermansiagydF4y2Ba种虫害在衰老小鼠结肠细胞腔的内容减少(gydF4y2Ba10gydF4y2Ba,gydF4y2Ba24gydF4y2Ba,gydF4y2Ba25gydF4y2Ba]。另一项研究也报道了一种与年龄相关的损失gydF4y2BaAkkermansiagydF4y2Ba种虫害在人类gydF4y2Ba26gydF4y2Ba]。有趣的是,大量的gydF4y2BaAkkermansiagydF4y2Ba种虫害是显示增加了百岁老人(105 - 109年)相比,年轻的年龄组(gydF4y2Ba27gydF4y2Ba]。这些结果可能表明老年之间存在关系达到一个极端的和丰富的gydF4y2BaAkkermansiagydF4y2Baspp。gydF4y2Ba24gydF4y2Ba,gydF4y2Ba27gydF4y2Ba]。gydF4y2Ba
许多潜在的有益的特点gydF4y2Ba答:muciniphilagydF4y2Ba表明,这种细菌微生物补充可能是一个强有力的候选人。然而,这种细菌对肠道健康的下降的影响而衰老期间还没有广泛调查。因此,本研究的目的是调查的影响,补充gydF4y2Ba答:muciniphilagydF4y2Ba对肠道健康的不同方面。我们使用gydF4y2BaErcc1gydF4y2Ba−/Δ7gydF4y2Ba老鼠,加速衰老小鼠模型,平均寿命~ 20周。进一步的特点这老鼠模型被广泛描述在先前的研究gydF4y2Ba28gydF4y2Ba,gydF4y2Ba29日gydF4y2Ba,gydF4y2Ba30.gydF4y2Ba),说明加速老化的表型gydF4y2BaErcc1gydF4y2Ba−/Δ7gydF4y2Ba老鼠在很大程度上类似于正常老化。的gydF4y2BaErcc1gydF4y2Ba−/Δ7gydF4y2Ba老鼠补充gydF4y2Ba答:muciniphilagydF4y2Ba通过口服填喂法为10周。牺牲后,回肠和结肠受到转录分析和微生物群组成在这些器官了。此外,我们评估粘液厚度在结肠癌和免疫细胞的分布的几种组织。gydF4y2Ba
结果gydF4y2Ba
答:muciniphilagydF4y2Ba补充的结肠粘液厚度增加gydF4y2BaErcc1gydF4y2Ba−/Δ7gydF4y2Ba老鼠gydF4y2Ba
自gydF4y2Ba答:muciniphilagydF4y2Bamucus-colonizing细菌,利用粘液作为能量来源,我们调查是否补充gydF4y2Ba答:muciniphilagydF4y2Ba在结肠黏液层产生影响的gydF4y2BaErcc1gydF4y2Ba−/Δ7gydF4y2Ba老鼠。粘液测量厚度的不是/阿尔新蓝染色结肠组织透露,老鼠的黏液层明显厚补充gydF4y2Ba答:muciniphilagydF4y2Ba相对于对照组(gydF4y2BapgydF4y2Ba< 0.001)(图gydF4y2Ba1gydF4y2Baa - c)。此外,结果与老鼠的粘液厚度补充gydF4y2Bal .杆菌gydF4y2Ba(WCFS1),因为我们之前显示,补充这种细菌阻止了与年龄相关的粘液厚度下降(gydF4y2Ba29日gydF4y2Ba]。结肠黏液层gydF4y2Bal .杆菌gydF4y2Ba老鼠是厚相比,对照组(补充gydF4y2BapgydF4y2Ba< 0.001)(图gydF4y2Ba1gydF4y2Baa, d),但补充gydF4y2Ba答:muciniphilagydF4y2Ba导致明显比厚粘液层gydF4y2Bal .杆菌gydF4y2Ba老鼠(补充gydF4y2BapgydF4y2Ba< 0.001)(图gydF4y2Ba1gydF4y2Ba)。这些结果表明,补充gydF4y2Ba答:muciniphilagydF4y2Ba有助于预防老年的黏液层厚度降低。gydF4y2Ba

粘液厚度增加gydF4y2BaErcc1gydF4y2Ba−/Δ7gydF4y2Ba老鼠补充gydF4y2BaAkkermansia muciniphilagydF4y2Ba。gydF4y2Ba一个gydF4y2Ba(μm)以结肠粘液厚度gydF4y2BaErcc1gydF4y2Ba−/Δ7gydF4y2Ba老鼠在控制,gydF4y2Ba答:muciniphilagydF4y2Ba集团和gydF4y2Bal .杆菌gydF4y2Ba组。gydF4y2BabgydF4y2Ba代表性的图片不是/阿尔新蓝染色控制鼠标,(gydF4y2BacgydF4y2Ba)鼠标补充gydF4y2Ba答:muciniphilagydF4y2Ba和(gydF4y2BadgydF4y2Ba)gydF4y2Bal .杆菌gydF4y2Ba。数据代表的意思是+扫描电镜从三到五个老鼠每组。* * *gydF4y2BapgydF4y2Ba< 0.001。μm规模酒吧组织学图片:100gydF4y2Ba
没有补充结肠和回肠后微生物群组成的差异gydF4y2Ba答:muciniphilagydF4y2Ba
为了调查是否与补充gydF4y2Ba答:muciniphilagydF4y2Ba引起的肠道微生物群组成的变化,我们16 s rRNA基因测序进行结肠和回肠内容。Alpha-diversity(香农熵),结肠和回肠内容样本没有控制和补充老鼠(图之间的不同。gydF4y2Ba2gydF4y2Baa)。调查是否与补充gydF4y2Ba答:muciniphilagydF4y2Ba导致这种细菌殖民化的增加,相对丰富的gydF4y2BaAkkermansiagydF4y2Ba种虫害在结肠内容进行评估。在结肠内容,干预组的平均相对丰度略高(0.738±1.279%)相比,对照组(0.252±0.503%)(图gydF4y2Ba2gydF4y2Bab),但这种差异没有统计学意义。gydF4y2BaAkkermansiagydF4y2Ba种虫害没有出现在回肠内容,除了一个鼠标在干预组(数据没有显示)。微生物组成样本之间的变化在主坐标分析(PCoA)基于Bray-Curtis不同。在结肠和回肠样本,没有明确的控制和之间的隔离观察gydF4y2Ba答:muciniphilagydF4y2Ba集团(无花果。gydF4y2Ba2gydF4y2Bac - d)。此外,在属水平没有明显的统计学差异之间的控制和补充老鼠被发现在两个来源(数据没有显示)。这些数据表明,细菌与补充gydF4y2Ba答:muciniphilagydF4y2Ba三次一个星期没有导致肠道微生物群组成的变化gydF4y2BaErcc1gydF4y2Ba−/Δ7gydF4y2Ba老鼠。gydF4y2Ba
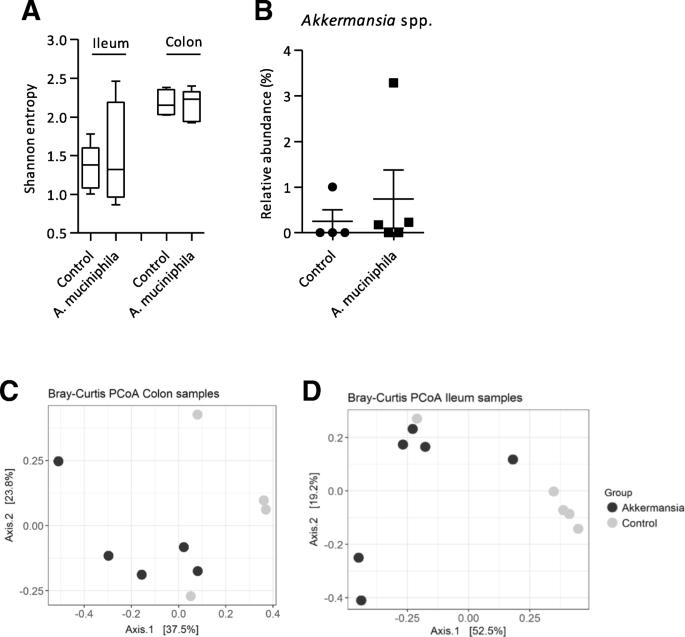
微生物群组成的结肠和回肠gydF4y2BaErcc1gydF4y2Ba−/Δ7gydF4y2Ba老鼠补充gydF4y2BaAkkermansia muciniphilagydF4y2Ba。gydF4y2Ba一个gydF4y2BaAlpha-diversity(香农熵)以回肠和结肠样本。框扩展从25到75,中线代表中间,胡须代表最小和最大值。gydF4y2BabgydF4y2Ba相对丰度(%)gydF4y2BaAkkermansiagydF4y2Ba种虫害在结肠内容评估16 s rRNA测序。数据代表的意思是+扫描电镜(gydF4y2BacgydF4y2Ba)Beta-diversity衡量Bray-Curtis结肠癌样本和主坐标分析(gydF4y2BadgydF4y2Ba)回肠样本。4和6之间每组小鼠被用于微生物群分析gydF4y2Ba
答:muciniphilagydF4y2Ba补充最低限度改变基因的表达在结肠肠屏障功能gydF4y2Ba
探索补充的影响gydF4y2Ba答:muciniphilagydF4y2Ba在基因表达,转录组分析在mRNA孤立的结肠和回肠组织gydF4y2BaErcc1gydF4y2Ba−/Δ7gydF4y2Ba老鼠。在结肠,数量427显著差异表达的基因(gydF4y2BapgydF4y2Ba1.2 < 0.05,褶皱变化>或<−1.2)之间的控制和gydF4y2Ba答:muciniphilagydF4y2Ba集团202年由225上调和下调的基因。从一个高度显著增加小鼠的结肠粘液厚度在接受gydF4y2Ba答:muciniphilagydF4y2Ba,结肠粘液生产调查相关基因的表达。没有显著差异的观察黏蛋白的表达,除了粘蛋白1 (gydF4y2BaMucl1gydF4y2Ba)这是下调(−1.5倍)gydF4y2Ba答:muciniphilagydF4y2Ba组相比,控制老鼠(额外的文件gydF4y2Ba1gydF4y2Ba)。此外,探索如果在结肠粘液厚度增加导致了增强肠道屏障功能,紧密连接功能相关的基因表达研究。经典的紧密连接蛋白gydF4y2BaTjp 1 - 3gydF4y2Ba,gydF4y2Ba果酱1 - 3gydF4y2Ba,gydF4y2BaClaudingydF4y2Ba家庭和gydF4y2BaOclngydF4y2Ba没有差异表达(数据没有显示)。调查在结肠途径被监管gydF4y2Ba答:muciniphilagydF4y2Ba补充,基因集富集分析(GSEA)。大大丰富了通路是由细胞周期相关流程,但是没有观察到通路相关的肠道屏障功能(附加文件gydF4y2Ba2gydF4y2Ba)。gydF4y2Ba
补充与gydF4y2Ba答:muciniphilagydF4y2Ba减少的表达基因和通路相关的抗菌活动、代谢过程和粘液回肠的生物合成gydF4y2Ba
自补充gydF4y2Ba答:muciniphilagydF4y2Ba最低限度改变相关的基因的表达在结肠肠屏障功能,我们也调查回肠组织的基因表达谱gydF4y2BaErcc1gydF4y2Ba−/Δ7gydF4y2Ba老鼠。795个基因的显著差异表达(gydF4y2BapgydF4y2Ba1.2 < 0.05,褶皱变化>或<−1.2)之间的控制和gydF4y2Ba答:muciniphilagydF4y2Ba集团在回肠,370年由425上调和下调的基因。有趣的是,多个基因编码的抗菌肽被抑制gydF4y2Ba答:muciniphilagydF4y2Ba补充的老鼠。gydF4y2BaReg3bgydF4y2Ba和gydF4y2BaReg3ggydF4y2Ba(无花果。gydF4y2Ba3gydF4y2Baa、b;额外的文件gydF4y2Ba3gydF4y2Ba)。然而,基因的表达编码alpha-defensins和溶菌酶不受影响gydF4y2Ba答:muciniphilagydF4y2Ba补充。紧密连接的差异表达基因被降到了最低程度,只有gydF4y2BaCldn2gydF4y2Ba和gydF4y2BaCldn8gydF4y2Ba被抑制在回肠的老鼠补充gydF4y2Ba答:muciniphilagydF4y2Ba而对照组(图。gydF4y2Ba3gydF4y2Bac, d;额外的文件gydF4y2Ba3gydF4y2Ba)。GSEA透露,大大丰富了抑制通路是由代谢过程(附加文件gydF4y2Ba2gydF4y2Ba)。此外,通路”N-Glycan生物合成”和“N-Glycan生物合成的前体(多萜醇Lipid-Linked低聚糖,LLO)和转移到新兴的蛋白质”被抑制在回肠的老鼠gydF4y2Ba答:muciniphilagydF4y2Ba补充相比对照组(附加文件gydF4y2Ba2gydF4y2Ba)。基于这一发现,微阵列数据集是寻找粘液生物合成相关的基因。的基因gydF4y2BaCtnna3gydF4y2Ba和gydF4y2BaSt6galnac6gydF4y2Ba被抑制在回肠补充小鼠和对照组(图。gydF4y2Ba3gydF4y2Bae, f;额外的文件gydF4y2Ba3gydF4y2Ba)。gydF4y2Ba
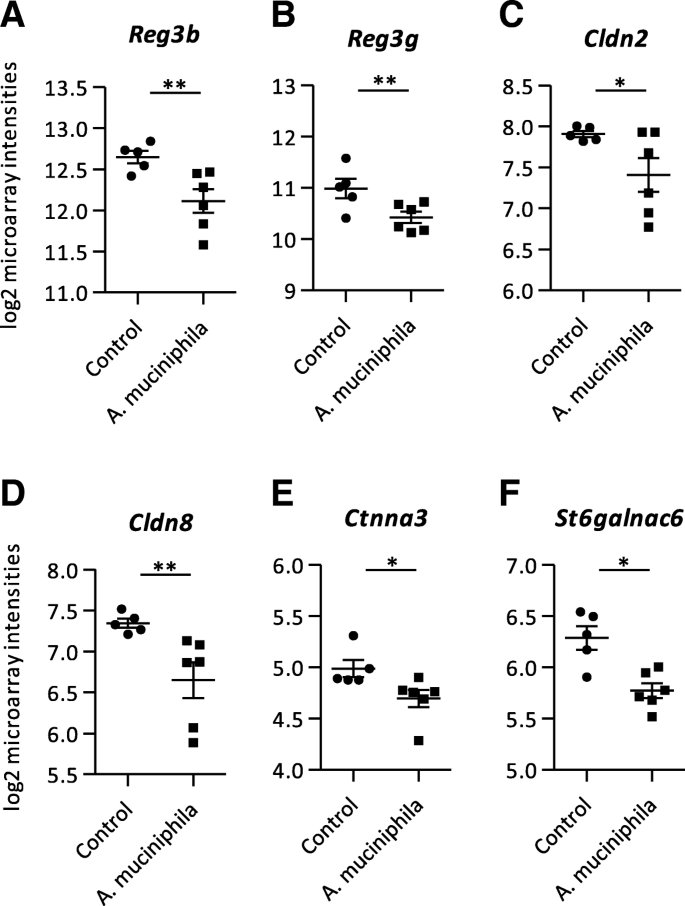
从回肠组织微阵列分析mRNA进行隔离。gydF4y2Ba一个gydF4y2BaLog2芯片强度的再生islet-derived 3β(gydF4y2BaReg3bgydF4y2Ba),(gydF4y2BabgydF4y2Ba3)再生islet-derivedγ(gydF4y2BaReg3ggydF4y2Ba)(gydF4y2BacgydF4y2Ba)Claudin 2 (gydF4y2BaCldn2gydF4y2Ba),(gydF4y2BadgydF4y2Ba)Claudin 8 (gydF4y2BaCldn8gydF4y2Ba),(gydF4y2BaegydF4y2Ba)连环蛋白(钙粘着蛋白相关蛋白),α3 (gydF4y2BaCtnna3gydF4y2Ba)和(gydF4y2BafgydF4y2Ba)ST6 (alpha-N-acetyl-neuraminyl-2 3-beta-galactosyl-1 3) -N-acetylgalactosaminideα2,6-sialyltransferase 6 (gydF4y2BaSt6galnac6gydF4y2Ba)。数据代表的意思是+ SEM。控制:gydF4y2BangydF4y2Ba= 5,gydF4y2Ba答:muciniphilagydF4y2Ba:gydF4y2BangydF4y2Ba= 6。*gydF4y2BapgydF4y2Ba< 0.05;* *gydF4y2BapgydF4y2Ba< 0.01gydF4y2Ba
减少表达与炎症和免疫功能相关的基因和通路在结肠和回肠gydF4y2Ba答:muciniphilagydF4y2Ba补充gydF4y2Ba
免疫功能是一个重要因素对肠道健康,免疫应答相关的基因表达谱。值得注意的是,抑制基因在结肠和回肠都由基因编码的免疫球蛋白(附加文件gydF4y2Ba1gydF4y2Ba)。趋化因子在结肠,多个基因编码等gydF4y2BaCxcl13gydF4y2Ba(无花果。gydF4y2Ba4gydF4y2Ba一个;额外的文件gydF4y2Ba3gydF4y2Ba),gydF4y2BaCcl12gydF4y2Ba,以及细胞因子gydF4y2BaIl5gydF4y2Ba和补充因素gydF4y2BaC1ragydF4y2Ba和gydF4y2BaC5ar1gydF4y2Ba都是下调(额外的文件吗gydF4y2Ba1gydF4y2Ba)。同时免疫球蛋白受体gydF4y2BaPilrb1gydF4y2Ba减少结肠的表达gydF4y2Ba答:muciniphilagydF4y2Ba集团(额外的文件gydF4y2Ba1gydF4y2Ba)。此外,其他的几种基因被抑制的结肠gydF4y2Ba答:muciniphilagydF4y2Ba补充与对照组相比,小鼠。gydF4y2Ba黑色gydF4y2Ba,gydF4y2BaCd4gydF4y2Ba,gydF4y2BaCd72gydF4y2Ba,gydF4y2BaTlr7gydF4y2Ba和gydF4y2BaTlr12gydF4y2Ba(无花果。gydF4y2Ba4gydF4y2Bab-f;额外的文件gydF4y2Ba3gydF4y2Ba)。GSEA显示的几种途径抑制在冒号,例如“肠道免疫网络IgA生产”,“Cytokine-Cytokine受体相互作用”和“炎症反应通路”,在别人(附加文件gydF4y2Ba2gydF4y2Ba)。此外,创造力通路分析(IPA)确定7细胞因子上游监管机构补充后将被抑制gydF4y2Ba答:muciniphilagydF4y2Ba,包括炎性摘要意思和抗炎及(表gydF4y2Ba1gydF4y2Ba)。还其他炎症相关的因素,如骨髓分化主要响应88 (Myd88)、肿瘤坏死因子受体超家族成员1 b (Tnfrsf1b), 12 (Tnfsf12) NFKBα抑制剂(Nfkbia), T细胞受体(TCR)和Toll样受体衔接分子1 (Ticam1)预测抑制上游监管者(表gydF4y2Ba1gydF4y2Ba)。在回肠,GSEA注释通路”抗原表示:折叠、装配和加载的类肽MHC”是高度上调(附加文件gydF4y2Ba2gydF4y2Ba)。最重要的上游调节器被异丙醇在回肠组织白介素10受体α亚基(Il10RA),预测后稍微抑制gydF4y2Ba答:muciniphilagydF4y2Ba补充(表gydF4y2Ba2gydF4y2Ba)。总的来说,这些结果表明,gydF4y2Ba答:muciniphilagydF4y2Ba补充减少许多基因和通路的表达与炎症和免疫功能在结肠和回肠。gydF4y2Ba
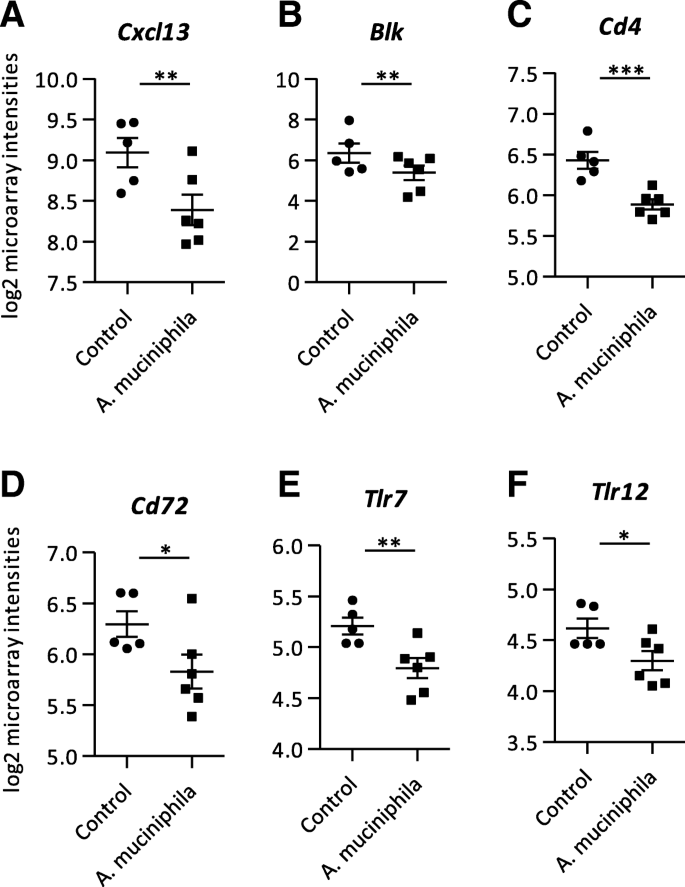
从结肠组织微阵列分析mRNA进行隔离。gydF4y2Ba一个gydF4y2BaLog2芯片强度(gydF4y2Ba一个gydF4y2Ba13)C-X-C主题趋化因子配体(gydF4y2BaCxcl13gydF4y2Ba),(gydF4y2BabgydF4y2Ba)B淋巴激酶(gydF4y2Ba黑色gydF4y2Ba),(gydF4y2BacgydF4y2Ba4)集群的区别(gydF4y2BaCd4gydF4y2Ba),(gydF4y2BadgydF4y2Ba72)集群的区别(gydF4y2BaCd72gydF4y2Ba),(gydF4y2BaegydF4y2Ba7)toll样受体(gydF4y2BaTlr7gydF4y2Ba),(gydF4y2BafgydF4y2Ba12)toll样受体(gydF4y2BaTlr12gydF4y2Ba)。数据代表的意思是+ SEM。对照组:gydF4y2BangydF4y2Ba= 5。gydF4y2Ba答:muciniphilagydF4y2Ba组:gydF4y2BangydF4y2Ba= 6。*gydF4y2BapgydF4y2Ba< 0.05;* *gydF4y2BapgydF4y2Ba< 0.01;* * *gydF4y2BapgydF4y2Ba< 0.001gydF4y2Ba
微小的变化在当地B细胞分布gydF4y2Ba答:muciniphilagydF4y2Ba补充gydF4y2Ba
基于微阵列分析的结果,我们继续调查免疫细胞分布在不同器官的免疫系统。因为几个B细胞相关基因被抑制gydF4y2Ba答:muciniphilagydF4y2Ba补充,如免疫球蛋白基因(回肠和结肠)gydF4y2Ba黑色gydF4y2Ba(只有在冒号),我们首先关注当地的B细胞的分布。肠系膜淋巴结(MLN)和派尔集合淋巴结补丁(PP),观察组之间没有差异B细胞频率(无花果。gydF4y2Ba5gydF4y2Baa - c)。然而,在PP激活CD80的频率gydF4y2Ba+gydF4y2BaCD273gydF4y2Ba−gydF4y2BaB细胞是补充显著降低小鼠(gydF4y2BapgydF4y2Ba= 0.009)(图gydF4y2Ba5gydF4y2Bad)。此外,频率的不成熟的CD80gydF4y2Ba−gydF4y2BaCD273gydF4y2Ba−gydF4y2Ba显著的高于B细胞gydF4y2Ba答:muciniphilagydF4y2Ba集团(gydF4y2BapgydF4y2Ba= 0.03)(图gydF4y2Ba5gydF4y2Bae),而没有CD80中观察到的变化gydF4y2Ba+gydF4y2BaCD273gydF4y2Ba+gydF4y2Ba存储器像和CD80gydF4y2Ba−gydF4y2BaCD273gydF4y2Ba+gydF4y2BaB细胞(图。gydF4y2Ba5gydF4y2Baf, g) (gydF4y2Ba31日gydF4y2Ba]。gydF4y2Ba

分布的B细胞子集肠系膜淋巴结(MLN)和派尔集合淋巴结补丁(PP)后补充gydF4y2BaAkkermansia muciniphilagydF4y2Ba。gydF4y2Ba一个gydF4y2Ba中频MLN的B细胞。gydF4y2BabgydF4y2Ba流仪结果页的B细胞。gydF4y2BacgydF4y2Ba意味着在PP B细胞的频率。gydF4y2BadgydF4y2Ba意味着CD80的频率gydF4y2Ba+gydF4y2BaCD273gydF4y2Ba−gydF4y2BaB细胞(gydF4y2BaegydF4y2Ba)CD80gydF4y2Ba−gydF4y2BaCD273gydF4y2Ba−gydF4y2BaB细胞(gydF4y2BafgydF4y2Ba)CD80gydF4y2Ba+gydF4y2BaCD273gydF4y2Ba+gydF4y2BaB细胞和gydF4y2BaggydF4y2BaCD80gydF4y2Ba−gydF4y2BaCD273gydF4y2Ba+gydF4y2BaB细胞数据代表的意思是+页。扫描电镜从5到6小鼠每组。*gydF4y2BapgydF4y2Ba< 0.05;* *gydF4y2BapgydF4y2Ba< 0.01gydF4y2Ba
答:muciniphilagydF4y2Ba补充增加脾脏B细胞的迁移和BMgydF4y2Ba
接下来,我们继续调查,B细胞在脾脏和骨髓(BM)子集。总在脾脏B细胞的频率并不是不同组(无花果。gydF4y2Ba6gydF4y2Ba)。其他在脾脏B细胞子集,如不成熟,滤泡和边缘区B细胞组之间没有显著差异(数据没有显示)。然而,B1细胞在脾脏的频率更高gydF4y2Ba答:muciniphilagydF4y2Ba老鼠(补充gydF4y2BapgydF4y2Ba= 0.02)(图gydF4y2Ba6gydF4y2Bab, c)。在骨髓(BM),更高频率的趋势的b细胞中观察到gydF4y2Ba答:muciniphilagydF4y2Ba补充的老鼠相比,控制老鼠(gydF4y2BapgydF4y2Ba= 0.07)和成熟和不成熟的B细胞的频率也更高(分别p = 0.02和0.06)(无花果。gydF4y2Ba6gydF4y2Bad)。频率的B细胞前体,即pro-B细胞,小休息前B细胞和大自行车前B细胞组没有差异(图gydF4y2Ba6gydF4y2Bah-j)。这些数据表明,gydF4y2Ba答:muciniphilagydF4y2Ba补充并没有改变生产新的B细胞在BM,但增加B细胞的迁移到脾脏和BM。gydF4y2Ba

B细胞亚型的分布在脾脏和骨髓(BM)补充gydF4y2BaAkkermansia muciniphilagydF4y2Ba。gydF4y2Ba一个gydF4y2Ba平均频率CD19gydF4y2Ba+gydF4y2BaCD3gydF4y2Ba−gydF4y2Ba在脾脏B细胞。gydF4y2BabgydF4y2Ba流仪分析单一CD19生活gydF4y2Ba+gydF4y2BaCD3gydF4y2Ba−gydF4y2Ba细胞在脾脏。gydF4y2BacgydF4y2Ba平均频率CD5gydF4y2Ba+gydF4y2BaCD43gydF4y2Ba+gydF4y2BaB1细胞在脾脏。gydF4y2BadgydF4y2Ba意味着在BM频率的B细胞。gydF4y2BaegydF4y2Ba流仪分析单一CD19生活gydF4y2Ba+gydF4y2Ba细胞,分为pro-B细胞,大的自行车前b细胞和小休息前b细胞在BM。gydF4y2BafgydF4y2Ba意味着成熟B细胞频率,gydF4y2BaggydF4y2Ba不成熟的B细胞,gydF4y2BahgydF4y2BaPro-B细胞,gydF4y2Ba我gydF4y2Ba小休息前b细胞(gydF4y2BajgydF4y2Ba在BM)大型自行车前b细胞。数据代表的意思是+扫描电镜从4到6个老鼠每组。*gydF4y2BapgydF4y2Ba< 0.05gydF4y2Ba
MLN和脾脏T细胞分布是不变的gydF4y2Ba答:muciniphilagydF4y2Ba补充gydF4y2Ba
转录组的分析表明,表达gydF4y2BaCd4gydF4y2Ba基因是减少小鼠的结肠gydF4y2Ba答:muciniphilagydF4y2Ba而控制老鼠。此外,音标显示T细胞受体(TCR)预测抑制上游调节器在结肠。因此,我们研究了T细胞的分布是否改变组脾脏和MLN之间。然而,CD4gydF4y2Ba+gydF4y2Ba和CD8gydF4y2Ba+gydF4y2BaT细胞分布在MLN和脾脏没有不同与对照组相比,无论是CD4细胞gydF4y2Ba+gydF4y2BaFoxP3gydF4y2Ba+gydF4y2Ba在这两种免疫组织(图Treg频率改变。gydF4y2Ba7gydF4y2Baf)。gydF4y2Ba

分布在肠系膜淋巴结T细胞(MLN)和脾脏后补充gydF4y2BaAkkermansia muciniphilagydF4y2Ba。gydF4y2Ba一个gydF4y2Ba是指CD4细胞的频率gydF4y2Ba+gydF4y2BaT细胞(gydF4y2BabgydF4y2Ba)CD8gydF4y2Ba+gydF4y2BaT细胞(gydF4y2BacgydF4y2Ba)具体gydF4y2Ba+gydF4y2Ba在MLN Treg细胞。gydF4y2BadgydF4y2Ba是指CD4细胞的频率gydF4y2Ba+gydF4y2BaT细胞(gydF4y2BaegydF4y2Ba)CD8gydF4y2Ba+gydF4y2BaT细胞(gydF4y2BafgydF4y2Ba)具体gydF4y2Ba+gydF4y2BaTreg细胞在脾脏。数据代表的意思是+扫描电镜从5到6小鼠每组gydF4y2Ba
减少炎症细胞群在脾脏和MLN之后gydF4y2Ba答:muciniphilagydF4y2Ba补充gydF4y2Ba
接下来,因为GSEA透露一个浓缩通路相关的炎症反应和免疫功能,我们调查是否炎性细胞频率改变后的补充gydF4y2BaErcc1gydF4y2Ba−/Δ7gydF4y2Ba老鼠。在脾脏,道达尔和Ly6C的频率gydF4y2Ba嗨gydF4y2Ba单核细胞中略低gydF4y2Ba答:muciniphilagydF4y2Ba集团和Ly6CgydF4y2BaintgydF4y2Ba单核细胞显著降低(gydF4y2BapgydF4y2Ba= 0.01)(图gydF4y2Ba8gydF4y2Ba模拟)。此外,中性粒细胞的频率也补充老鼠(图略低。gydF4y2Ba8gydF4y2Bae)。同样的趋势这些炎性细胞群被发现在MLN,尽管细胞频率较低(< 1%,数据未显示)。这些数据显示出轻微的减少在脾脏和MLN的炎症标记物gydF4y2Ba答:muciniphilagydF4y2Ba补充在gydF4y2BaErcc1gydF4y2Ba−/Δ7gydF4y2Ba老鼠。此外,我们评估我们是否可以识别任何免疫细胞渗透的迹象通过调查)染色的组织。结肠和回肠组织,没有组织免疫细胞渗透的迹象在控制和观察gydF4y2Ba答:muciniphilagydF4y2Ba集团(无花果。gydF4y2Ba8gydF4y2Baf)。gydF4y2Ba

在脾脏炎性细胞的分布频率补充gydF4y2BaAkkermansia muciniphilagydF4y2Ba。gydF4y2Ba一个gydF4y2Ba的意思是总CD11b的频率gydF4y2Ba+gydF4y2BaCD68gydF4y2Ba+gydF4y2Ba单核细胞。gydF4y2BabgydF4y2Ba流仪分析单一CD11b生活gydF4y2Ba+gydF4y2Ba在Ly6C Ly6C / CD68细胞分裂gydF4y2Ba嗨gydF4y2Ba,Ly6CgydF4y2BaintgydF4y2Ba,Ly6CgydF4y2Ba罗gydF4y2Ba单核细胞和CD68gydF4y2Ba昏暗的gydF4y2BaLy6CgydF4y2Baint /嗨gydF4y2Ba中性粒细胞。gydF4y2BacgydF4y2Ba平均频率Ly6CgydF4y2Ba嗨gydF4y2Ba单核细胞(gydF4y2BadgydF4y2Ba)Ly6CgydF4y2BaintgydF4y2Ba单核细胞(gydF4y2BaegydF4y2Ba中性粒细胞。(gydF4y2BafgydF4y2Ba)代表的照片)彩色结肠和回肠组织控制,gydF4y2Ba答:muciniphilagydF4y2Ba补充老鼠。数据代表的意思是+扫描电镜从5到6小鼠每组。*gydF4y2BapgydF4y2Ba< 0.05gydF4y2Ba
增加数量的CD11bgydF4y2Ba+gydF4y2Ba细胞和居民的腹膜巨噬细胞gydF4y2BaErcc1gydF4y2Ba−/Δ7gydF4y2Ba老鼠补充gydF4y2Ba答:muciniphilagydF4y2Ba
因为我们发现,大量的B1细胞在脾脏和细胞类型通常是高纯度的腹膜腔,我们还在腹膜免疫细胞的分布进行了调查。值得注意的是,在小鼠腹膜细胞补充的总数gydF4y2Ba答:muciniphilagydF4y2Ba几乎是三倍高于控制老鼠(gydF4y2BapgydF4y2Ba= 0.02)(图gydF4y2Ba9gydF4y2Baa)。B细胞的绝对数字,B1细胞和T细胞组之间没有显著差异(图gydF4y2Ba9gydF4y2Ba罪犯)。我们发现CD11b的绝对数量的增加gydF4y2Ba+gydF4y2Ba细胞在腹膜后gydF4y2Ba答:muciniphilagydF4y2Ba补充(gydF4y2BapgydF4y2Ba= 0.004)(图gydF4y2Ba9gydF4y2Ba调查CD11b e)gydF4y2Ba+gydF4y2Ba细胞子集透露,绝对中性粒细胞的数量没有显著更高gydF4y2Ba答:muciniphilagydF4y2Ba补充(无花果。gydF4y2Ba9gydF4y2Ba相比之下常驻巨噬细胞(f)gydF4y2BapgydF4y2Ba= 0.045)(图gydF4y2Ba9gydF4y2Bag h)。此外,CD115常驻巨噬细胞上的表达明显高于gydF4y2Ba答:muciniphilagydF4y2Ba组相对于对照组(gydF4y2BapgydF4y2Ba= 0.02)(图gydF4y2Ba9gydF4y2Ba我),而没有改变CD11b的表达和SIRPα观察(图。gydF4y2Ba9gydF4y2Baj, k)。gydF4y2Ba

免疫细胞分布在腹膜后补充gydF4y2BaAkkermansia muciniphilagydF4y2Ba。gydF4y2Ba一个gydF4y2Ba绝对数量总腹膜渗出物的细胞(压电陶瓷),(gydF4y2BabgydF4y2Ba)CD3gydF4y2Ba−gydF4y2BaCD19gydF4y2Ba+gydF4y2BaB细胞(gydF4y2BacgydF4y2Ba)CD5gydF4y2Ba+gydF4y2BaCD43gydF4y2Ba+gydF4y2BaB1细胞(gydF4y2BadgydF4y2Ba)CD3gydF4y2Ba+gydF4y2BaT细胞(gydF4y2BaegydF4y2Ba)CD11bgydF4y2Ba+gydF4y2Ba细胞和(gydF4y2BafgydF4y2Ba)CD11bgydF4y2Ba+gydF4y2BaLy6GgydF4y2Ba+gydF4y2Ba中性粒细胞。gydF4y2BaggydF4y2Ba流仪CD11b分析gydF4y2Ba+gydF4y2Ba细胞。gydF4y2BahgydF4y2Ba绝对数量的CD11bgydF4y2Ba+gydF4y2BaLy6GgydF4y2Ba−gydF4y2Ba居民在腹膜巨噬细胞渗出物细胞悬液。gydF4y2Ba我gydF4y2Ba平均荧光强度(MFI) CD115标记,(gydF4y2BajgydF4y2Ba)CD11b标记和(gydF4y2BakgydF4y2Ba常驻巨噬细胞)SIRPa标记表示。数据代表的意思是+扫描电镜从5到6小鼠每组。*gydF4y2BapgydF4y2Ba< 0.05;* *gydF4y2BapgydF4y2Ba< 0.01gydF4y2Ba
答:muciniphilagydF4y2Ba补充并没有改变生存,体重和器官的重量gydF4y2BaErcc1gydF4y2Ba−/Δ7gydF4y2Ba老鼠gydF4y2Ba
补充与gydF4y2Ba答:muciniphilagydF4y2Ba10周没有改变存活率比控制老鼠(额外的文件gydF4y2Ba4gydF4y2Ba)。身体的重量gydF4y2BaErcc1gydF4y2Ba−/Δ7gydF4y2Ba老鼠增加了上半年的生活,但又减少从11周大开始(附加文件gydF4y2Ba4gydF4y2Ba)。没有发现明显的体重差异发展团体之间。的重量肝脏,脾脏和胸腺测量后直接牺牲之间的控制和没有显著不同gydF4y2Ba答:muciniphilagydF4y2Ba(数据未显示)。gydF4y2Ba
讨论gydF4y2Ba
细菌gydF4y2BaAkkermansia muciniphilagydF4y2Ba建议是一种很有前途的微生物补充,由于其有益的影响(gydF4y2Ba20.gydF4y2Ba,gydF4y2Ba32gydF4y2Ba]。然而,它的影响肠道健康的下降在老化还没有好了。在目前的研究中,我们调查的影响,补充gydF4y2Ba答:muciniphilagydF4y2Ba在加速老化的肠道健康的不同方面gydF4y2BaErcc1gydF4y2Ba−/Δ7gydF4y2Ba老鼠。我们报告的补充gydF4y2Ba答:muciniphilagydF4y2Ba10周导致明显厚结肠黏液层和抗炎免疫状态的改善与对照组相比。gydF4y2Ba
在最近的两项研究,它表明降低衰老小鼠结肠黏液层,建议协会与细菌渗透和免疫激活gydF4y2Ba29日gydF4y2Ba,gydF4y2Ba33gydF4y2Ba]。一个体内减少粘液的厚度也观察到gydF4y2BaErcc1gydF4y2Ba−/Δ7gydF4y2Ba老鼠(gydF4y2Ba29日gydF4y2Ba),确认在衰老表型相似性在这个加速老化小鼠模型相比,小鼠模型和正常的衰老过程。我们之前显示,gydF4y2Bal .杆菌gydF4y2BaWCFS1结肠黏液层的厚度增加gydF4y2BaErcc1gydF4y2Ba−/Δ7gydF4y2Ba老鼠(gydF4y2Ba29日gydF4y2Ba),但我们现在显示gydF4y2Ba答:muciniphilagydF4y2Ba能进一步勾芡黏液层。的能力gydF4y2Ba答:muciniphilagydF4y2Ba增加粘液厚度也报道之前(gydF4y2Ba23gydF4y2Ba]。gydF4y2Ba答:muciniphilagydF4y2Ba能够降低粘蛋白结构使用它作为碳和氮源,因此称为mucus-degrader [gydF4y2Ba17gydF4y2Ba]。然而,观察到的黏液层厚度增加补充后表明,这种细菌能够积极打开主机结肠粘液生产,建议也一直由Derrien和他的同事们(gydF4y2Ba34gydF4y2Ba]。有趣的是,我们没有找到任何黏蛋白基因编码的微分表达式,除了下调gydF4y2BaMucl1gydF4y2Ba在结肠癌组织中。然而,这种基因的表达水平很低,它不是一个典型的粘蛋白,构成了结肠黏液层等gydF4y2BaMuc2gydF4y2Ba(gydF4y2Ba35gydF4y2Ba]。在我们之前的研究中,补充gydF4y2Bal .杆菌gydF4y2BaWCFS1并没有导致粘蛋白基因表达的差异,而显著增加观察结肠粘液厚度(gydF4y2Ba29日gydF4y2Ba]。可能,补充gydF4y2Ba答:muciniphilagydF4y2Ba导致了粘液厚度增加了影响粘液而不影响生物合成过程gydF4y2BaMuc2gydF4y2Ba表达式。为了了解确切的粘液流动过程的潜在机制,进一步调查是十分必要的。gydF4y2Ba
有趣的是,在回肠gydF4y2Ba答:muciniphilagydF4y2Ba补充老鼠,GSEA透露两个N-Glycan生物合成途径相关的下调,除此之外,两个粘液生物合成相关的基因也被下调。这些结果暗示,除了其对结肠黏液层产生巨大影响,gydF4y2Ba答:muciniphilagydF4y2Ba也可以在回肠粘膜层产生影响。我们无法验证这一发现通过测量回肠粘膜层,因为它而不连续出现由于绒毛的存在。gydF4y2Ba
黏液层的重要功能,紧密连接密封套肠道上皮细胞中也扮演着重要的角色在肠屏障功能和TJ的年龄相关性降低表达基因被发现在狒狒gydF4y2Ba36gydF4y2Ba]。我们和其他人之前显示gydF4y2Ba答:muciniphilagydF4y2Ba改善肠屏障功能Caco-2细胞模型(gydF4y2Ba37gydF4y2Ba)以及小鼠(gydF4y2Ba18gydF4y2Ba,gydF4y2Ba19gydF4y2Ba,gydF4y2Ba23gydF4y2Ba,gydF4y2Ba38gydF4y2Ba,gydF4y2Ba39gydF4y2Ba]。在目前的研究中,我们发现的下调gydF4y2BaCldn2gydF4y2Ba和gydF4y2BaCldn8gydF4y2Ba回肠的gydF4y2Ba答:muciniphilagydF4y2Ba补充老鼠。然而,肠道屏障由多个蛋白质网络的复杂结构gydF4y2Ba40gydF4y2Ba]。因此,它是不可能任何固体得出结论的影响gydF4y2Ba答:muciniphilagydF4y2Ba补充对肠道屏障功能基于基因表达数据。gydF4y2Ba
肠道微生物群组成和细菌多样性回肠和结肠补充后没有显著改变gydF4y2Ba答:muciniphilagydF4y2Ba。这一结果表明,细菌与补充gydF4y2Ba答:muciniphilagydF4y2Ba不会导致一个重塑的肠道微生物群组成,这也是之前报道(gydF4y2Ba23gydF4y2Ba]。肠道微生物群的分析显示的相对丰度gydF4y2Ba答:muciniphilagydF4y2Ba在回肠和结肠内容低。这是之前显示gydF4y2BaAkkermansiagydF4y2Ba种虫害在mono-colonised老鼠比回肠结肠中(gydF4y2Ba39gydF4y2Ba),但在结肠低丰度显著。剂量的细菌,即2×10gydF4y2Ba8gydF4y2BaCFU一周3次,在先前的老鼠研究(已被证明有效gydF4y2Ba23gydF4y2Ba]。可能最后口头填喂法之间的相对较长的时间和牺牲(约24小时)可能导致的冲刷gydF4y2Ba答:muciniphilagydF4y2Ba。不过,最近的一项研究表明,每天补充11益生菌菌株导致低殖民在小鼠造成的土著微生物(gydF4y2Ba41gydF4y2Ba]。也许,这一发现也可以解释了阻碍了殖民统治gydF4y2Ba答:muciniphilagydF4y2Ba在我们的研究中。gydF4y2Ba
细菌与补充gydF4y2Ba答:muciniphilagydF4y2Ba导致大量的免疫相关基因的下调,在结肠途径。值得注意的是,这些包括几个B细胞相关基因,如免疫球蛋白、gydF4y2Ba黑色gydF4y2Ba和gydF4y2BaPilrb1gydF4y2Ba,在别人。此外,GSEA透露的下调途径“肠道免疫网络IgA生产”。这些结果暗示gydF4y2Ba答:muciniphilagydF4y2Ba补充可能降低生产IgA的必要性,即施加粘膜保护反应对共生的细菌(gydF4y2Ba42gydF4y2Ba]。根据这些结果,我们发现两者的下调gydF4y2BaTlr7gydF4y2Ba和gydF4y2BaTlr12gydF4y2Ba在结肠。然而,我们无法证实这一假设,因为IgA浓度在结肠没有测量在这个研究。我们做了调查B细胞频率在几个免疫组织。然而,我们不能找到差异在mln的B细胞频率,而B细胞在脾脏和BM频率增加。这些研究结果表明,基于转录组分析,gydF4y2Ba答:muciniphilagydF4y2BaB细胞频率和CD4下降gydF4y2Ba+gydF4y2Ba导致结肠癌和T细胞的潜在再分配B和T细胞在淋巴器官。相反,总在BM和脾脏B细胞子集的频率略有增加gydF4y2Ba答:muciniphilagydF4y2Ba补充,没有在BM B细胞前体的变化频率。这些发现可能表明补充gydF4y2Ba答:muciniphilagydF4y2Ba抑制涌入的B细胞进入结肠,导致稍微增加成熟B细胞池在脾脏和BM。gydF4y2Ba
此外,我们发现在结肠炎症标记物减少gydF4y2Ba答:muciniphilagydF4y2Ba补充。几个基因编码的趋化因子,补充因素,以及细胞因子gydF4y2BaIl5gydF4y2Ba后被抑制gydF4y2Ba答:muciniphilagydF4y2Ba补充。此外,异丙醇认为大量的促炎细胞因子可能抑制上游结肠的监管机构。的抗炎作用gydF4y2Ba答:muciniphilagydF4y2Ba已经广泛的描述(gydF4y2Ba34gydF4y2Ba]。我们现在增加证据gydF4y2Ba答:muciniphilagydF4y2Ba可以防止体内增加的炎症(inflamm-aging)通过减少结肠炎性基因和通路的表情。此外,我们还发现了抗炎细胞因子及抑制上游调节器。虽然我们只发现一个抗炎细胞因子,这一发现可能指向一个通用减少免疫激活的gydF4y2Ba答:muciniphilagydF4y2Ba。组织学分析结肠并没有透露任何明确两组免疫渗透的迹象。Derrien和他的同事在此前的一项研究中的免疫相关基因的上调与没有显微镜下可见的迹象炎症组织中也观察到小鼠mono-colonizedgydF4y2Ba答:muciniphilagydF4y2Ba(gydF4y2Ba39gydF4y2Ba]。作者认为这些观察免疫耐受对监管过程的一部分gydF4y2Ba答:muciniphilagydF4y2Ba。然而,在这项研究中使用无菌鼠,因此任何对比这些特定的结果和我们的结果应该谨慎。gydF4y2Ba
有趣的是,我们发现显著降低频率的激活B细胞亚型和更高的频率更成熟B细胞亚型在页的增加水平不成熟B细胞是按照从回肠组织微阵列的结果,因为我们还发现许多immunoglobulin-related基因的表达减少。除此之外,gydF4y2BaReg3bgydF4y2Ba和gydF4y2BaReg3ggydF4y2Ba都是抑制在回肠。我们之前显示,gydF4y2Ba答:muciniphilagydF4y2Ba补充也减少了gydF4y2BaReg3ggydF4y2Ba表达在回肠的高脂肪饮食的老鼠gydF4y2Ba19gydF4y2Ba]。在最近的一项研究中,增加抗菌基因的表达被发现回肠的C57BL / 6小鼠,其中包括gydF4y2BaReg3bgydF4y2Ba,gydF4y2BaReg3ggydF4y2Ba,gydF4y2BaDefb1gydF4y2Ba和gydF4y2BaRetnlbgydF4y2Ba,建议与增加上皮窘迫的状态(gydF4y2Ba43gydF4y2Ba]。因此,基于我们的研究结果我们建议补充gydF4y2Ba答:muciniphilagydF4y2Ba可能有助于预防与年龄相关的上皮在回肠痛苦。gydF4y2Ba
众所周知,在老化过程中T细胞功能下降(gydF4y2Ba44gydF4y2Ba]。我们先前的研究显示,补充gydF4y2Bal .杆菌gydF4y2BaWCFS1和gydF4y2Bal .干酪乳杆菌gydF4y2BaBL23 MLN的调节性T细胞的频率增加gydF4y2BaErcc1gydF4y2Ba−/Δ7gydF4y2Ba老鼠。然而,补充gydF4y2Ba答:muciniphilagydF4y2Ba没有导致任何改变在MLN的T细胞分布,脾和PP、尽管下调的吗gydF4y2BaCd4gydF4y2Ba基因的预测抑制结肠癌和上游监管机构识别。可能增加结肠黏液层所致gydF4y2Ba答:muciniphilagydF4y2Ba补充导致增加在结肠中保护状态,从而减少生产的T细胞吸引趋化因子和随后导致CD4的减少gydF4y2Ba+gydF4y2BaT细胞吸引和渗透。gydF4y2Ba
在腹腔,高度在居民观察巨噬细胞显著增加gydF4y2Ba答:muciniphilagydF4y2Ba补充。腹膜巨噬细胞是免疫反应的重要的调制在感染和促进组织内稳态(gydF4y2Ba45gydF4y2Ba]。此外,腹膜巨噬细胞被证明居民抵御微生物入侵(gydF4y2Ba46gydF4y2Ba),这可以解释高的细胞类型后补充gydF4y2Ba答:muciniphilagydF4y2Ba。然而,这增加居民腹膜巨噬细胞并不是伴随着增加频率的中性粒细胞和T细胞,而这些细胞类型期间预计将高度存在炎症。这个观察意味着补充gydF4y2Ba答:muciniphilagydF4y2Ba导致了激活状态对腹膜巨噬细胞,但导致了抗炎而非炎性反应。gydF4y2Ba
结论gydF4y2Ba
的关注gydF4y2Ba答:muciniphilagydF4y2Ba作为一个潜在的微生物补充增加了和充足的证据强调这种细菌的有利影响低度炎症和(有氧运动)代谢紊乱gydF4y2Ba16gydF4y2Ba,gydF4y2Ba20.gydF4y2Ba,gydF4y2Ba34gydF4y2Ba,gydF4y2Ba47gydF4y2Ba]。在目前的研究中,我们也发现一些在回肠代谢过程,以及在回肠和结肠都受到免疫过程gydF4y2Ba答:muciniphilagydF4y2Ba,但现在在老龄化的模型。此外,我们令人信服地表明,gydF4y2Ba答:muciniphilagydF4y2Ba有保护作用与年龄相关的粘液厚度下降,这是相比更强gydF4y2Bal .杆菌gydF4y2BaWCFS1。衰老往往伴随着慢性炎症和代谢综合征风险增加(gydF4y2Ba1gydF4y2Ba,gydF4y2Ba48gydF4y2Ba),导致生活质量下降和相当大的医疗成本上升gydF4y2Ba49gydF4y2Ba]。我们的研究显示之间的因果关系gydF4y2Ba答:muciniphilagydF4y2Ba和衰减的衰老表型,防止老年性结肠黏液层的厚度下降,提高免疫功能。因此,这些结果支持治疗的应用gydF4y2Ba答:muciniphilagydF4y2Ba在人口老龄化和调查为进一步研究铺平道路gydF4y2Ba答:muciniphilagydF4y2Ba作为治疗干预导致健康老龄化。进一步的研究应该集中在实用方面应用在人类身上,如剂量、频率和管理的方式。gydF4y2Ba
材料和方法gydF4y2Ba
老鼠和研究设计gydF4y2Ba
在这项研究中,加速老化gydF4y2BaErcc1gydF4y2Ba−/Δ7gydF4y2Ba老鼠使用。的基因的小鼠模型是广泛被别人前面描述的gydF4y2Ba28gydF4y2Ba,gydF4y2Ba29日gydF4y2Ba,gydF4y2Ba30.gydF4y2Ba]。简而言之,gydF4y2BaErcc1gydF4y2Ba−/Δ7gydF4y2Ba老鼠有一个受损的DNA修复蛋白ERCC1,导致积累多种DNA损伤,从而加速老化。老鼠分别安置在防晒系数条件下,收到一份随意纯化饮食(D12450B公式,研究饮食,附加文件gydF4y2Ba5gydF4y2Ba),并随意访问水由水瓶长喷嘴。老鼠补充gydF4y2BaAkkermansia muciniphilagydF4y2Ba民大gydF4y2BaTgydF4y2Ba(ATTC baa - 835)的口服剂量的2×10强饲法gydF4y2Ba8gydF4y2Ba200 CFU /μL,每周3次,总共10周。早上口语强饲法给出了。对照组同时收到口头填喂法包含相同体积的PBS。第三个gydF4y2BaErcc1gydF4y2Ba−/Δ7gydF4y2Ba小鼠组包括接受相同的剂量gydF4y2Ba乳杆菌gydF4y2BaWCFS1。这些老鼠只用于组织学的目的。细菌培养的生长过程是广泛前面描述的gydF4y2Ba29日gydF4y2Ba]。每组18老鼠的数量(男性和女性)包括,这些老鼠的寿命评估。10周后,当老鼠16周,5 - 6雌性老鼠被牺牲掉了。结肠和回肠内容,以及回肠末端和近端结肠部分,收集和snap-frozen液氮。一段回肠和结肠组织固定在Carnoy组织学目的的解决方案。脾脏,肠系膜淋巴结,派尔集合淋巴结补丁、骨髓和腹膜渗出物为免疫细胞被孤立的测量。gydF4y2Ba
组织学gydF4y2Ba
石蜡包埋后,Carnoy-fixed回肠末端和近端结肠组织切片在5μM部分poly-l-lysine镀膜玻璃幻灯片(热科学、德国)。幻灯片是脱蜡、脱水和苏木精和伊红染色())和私人助理/阿尔新蓝。结肠黏液层的厚度测量使用ImageJ软件(美国国立卫生研究院,医学博士)。比较黏液层的厚度,我们包括一个额外的鼠标组接受gydF4y2Ba乳杆菌gydF4y2BaWCFS1。gydF4y2Ba
微生物群成分分析gydF4y2Ba
DNA与回肠和结肠内容使用修改后的重复珠打方法(gydF4y2Ba50gydF4y2Ba]。微生物群组成评估MiSeq平台上使用16 s rRNA测序(Illumina公司、圣地亚哥、钙、美国)。接下来,NG-Tax管道是用于barcode-primer过滤,de-multiplexing, OTU和分类学分类(gydF4y2Ba51gydF4y2Ba]。生成的biom-files用于总结微生物数据,即alpha-diversity beta-diversity,使用r程序微生物[gydF4y2Ba52gydF4y2Ba]和phyloseq [gydF4y2Ba53gydF4y2Ba]。gydF4y2Ba
RNA隔离gydF4y2Ba
RNA是孤立的回肠末端和近端结肠组织(gydF4y2BangydF4y2Ba= 5 - 6小鼠/组)使用试剂盒试剂(表达载体、布雷达,荷兰)。净化的孤立的RNA进行使用RNeasy迷你包(Venlo试剂盒,荷兰)。测量总RNA的收益率(Nanodrop后,nd - 1000, Nanodrop产品,Maarssen,荷兰),RNA完整性评估(安捷伦2100生物分析仪,安捷伦科技,阿姆斯特丹,荷兰)和只包含RNA的RNA完整数量(RIN) 8.0以上。gydF4y2Ba
微阵列分析gydF4y2Ba
微阵列分析如前所述[gydF4y2Ba54gydF4y2Ba]。控制和之间的基因表达差异gydF4y2Ba答:muciniphilagydF4y2Ba补充组小鼠评估使用基于强度的主持T统计(简称IBMT)方法,gydF4y2BapgydF4y2Ba值< 0.05和<−1.2或> 1.2折变化。在NCBI的基因表达微阵列数据已经提交给综合(GEO) (GSE126730)。基因集富集分析(GSEA)是用来确定显著富集通路(gydF4y2Ba55gydF4y2Ba]。只有通路错误发现率(罗斯福)< 0.2考虑在内。创新路径分析(IPA)被用于识别上游监管机构(gydF4y2Ba56gydF4y2Ba]。gydF4y2Ba
互补脱氧核糖核酸合成和实时定量PCRgydF4y2Ba
实时定量PCR (qPCR)是用于验证差异表达的基因的表达谱芯片中确认分析。结肠和回肠样本,互补DNA合成(互补)从1000年使用RevertAid ng总RNA的合成第一链cDNA工具包(热费希尔科学、Landsmeer、荷兰)后,制造商的协议。以下使用热循环条件:5分钟25°C, 60分钟37°C和5分钟在70°C。引物序列得到在线PrimerBank数据库(附加文件gydF4y2Ba5gydF4y2Bab) (gydF4y2Ba57gydF4y2Ba]。执行qPCR CFX384热循环(Bio-Rad实验室、Veenendaal、荷兰)使用SensiMix SYBR No-ROX工具包(位于莱茵河畔Bioline,荷兰)。管家基因36 b4用于规范化。gydF4y2Ba
Fluorescence-activated细胞排序(流式细胞仪)分析gydF4y2Ba
脾脏,肠系膜淋巴结,派尔集合淋巴结补丁、骨髓和腹膜渗出物细胞都受制于流式细胞仪分析,类似如前所报道(gydF4y2Ba29日gydF4y2Ba]。总之,股骨,胫骨,回肠的波峰,前腿,胸骨收获,用研钵和研杵碎。Singe-cell悬浮液从每个器官都由细胞穿过40-μm细胞用注射器过滤器。首先,细胞被染色的细胞外的标记。可固定的生活/死eFluor506染色(美国圣地亚哥Ebioscience)是用来排除死细胞。细胞被固定和permeabilized修复/烫缓冲区(Ebioscience)为细胞内染色标记。所有抗体用于流式细胞术都加入了额外的文件gydF4y2Ba5gydF4y2Bac。第二章使用流式细胞分析仪(BD生物科学、Erembodegem、比利时)和数据分析使用FlowJo vX。07软件(树明星Inc .)、美国)。gydF4y2Ba
统计分析gydF4y2Ba
Kolmogorov-Smirnov测试用于测试数据是正态分布和适当的非参数统计检验时使用数据不是正态分布。对生存分析,log-rank (Mantel-Cox)测试使用。测试控制和补充组之间的差异,学生学习任务或Mann-Whitney U测试。关于粘液厚度的差异,克鲁斯卡尔-沃利斯检验邓恩的多个对比测试了,因为三个老鼠组。除非另有说明,gydF4y2BapgydF4y2Ba值的水平gydF4y2BapgydF4y2Ba< 0.05被认为是具有统计学意义。gydF4y2Ba
缩写gydF4y2Ba
- BM:gydF4y2Ba
-
骨髓gydF4y2Ba
- 菌落:gydF4y2Ba
-
克隆形成单位gydF4y2Ba
- Ercc1gydF4y2Ba——/Δ7gydF4y2Ba老鼠:gydF4y2Ba
-
核苷酸切除修复基因Ercc1的小老鼠gydF4y2Ba
- 流式细胞仪:gydF4y2Ba
-
Fluorescence-activated细胞分类gydF4y2Ba
- 舰队指挥官:gydF4y2Ba
-
褶皱变化gydF4y2Ba
- 罗斯福:gydF4y2Ba
-
错误发现率gydF4y2Ba
- GSEA:gydF4y2Ba
-
基因集富集分析gydF4y2Ba
- ):gydF4y2Ba
-
苏木精和伊红gydF4y2Ba
- 共同参与:gydF4y2Ba
-
基于强度的主持T统计数据gydF4y2Ba
- 异丙醇:gydF4y2Ba
-
创新途径分析gydF4y2Ba
- MLN:gydF4y2Ba
-
肠系膜淋巴结gydF4y2Ba
- 不是:gydF4y2Ba
-
周期性acid-SchiffgydF4y2Ba
- PCoA:gydF4y2Ba
-
主坐标分析gydF4y2Ba
- 页:gydF4y2Ba
-
淋巴集结gydF4y2Ba
- qPCR:gydF4y2Ba
-
定量聚合酶链反应gydF4y2Ba
- 防晒系数:gydF4y2Ba
-
特定的无菌gydF4y2Ba
- TJ:gydF4y2Ba
-
紧密连接gydF4y2Ba
引用gydF4y2Ba
森Bektas A》SH R,费鲁奇l .老化,炎症和环境。Exp Gerontol。2018; 105:10-8。gydF4y2Ba
布福德太瓦。(Dis)信任你的直觉:肠道微生物组在年龄相关的炎症,健康和疾病。微生物。2017;5 (1):80。gydF4y2Ba
Sanada F, Taniyama Y, Muratsu J,大津R,清水H, Rakugi H,等。慢性炎症在衰老的来源。前面心血管医学。2018;12。gydF4y2Ba
Franceschi C, Campisi j .慢性炎症(inflammaging)及其潜在的贡献与年龄有关的疾病。J Gerontol Sci地中海Sci杂志。2014;69 (1):S4-9。gydF4y2Ba
雷纳CK,霍洛维茨m .生理学的肠道老化。当前的临床营养与代谢护理。2013;16 (1):33 - 8。gydF4y2Ba
Koh, De vad F, Kovatcheva-Datchary P, Backhed F从膳食纤维到主机生理学:短链脂肪酸的主要细菌代谢物。细胞。2016;165 (6):1332 - 45。gydF4y2Ba
路易斯·P, GL,弗林特HJ。肠道微生物群,细菌代谢物和结肠直肠癌。Nat Microbiol牧师。2014;12 (10):661 - 72。gydF4y2Ba
Belkaid Y,手太瓦。微生物群的免疫和炎症的作用。细胞。2014;157 (1):121 - 41。gydF4y2Ba
比如C,布列塔尼的J,丹尼尔·C Foligne b .维护肠道健康的生态系统:暂时的食品bug偷渡者和船员的一部分吗?食品Microbiol Int J。2015; 213:139-43。gydF4y2Ba
Fransen F,凡发现AA, Borghuis T,之后,Hugenholtz F, van der Gaast-de Jongh C, et al。年龄在肠道微生物群有助于Systemical Inflammaging后转移到无菌鼠。Immunol前面。2017;8:1385。gydF4y2Ba
McGuckin妈,林登SK,萨顿P弗罗林TH。粘蛋白动力学和肠道病原体。Nat Microbiol牧师。2011;9 (4):265 - 78。gydF4y2Ba
Van der Sluis M, De通力英航De Bruijn AC Velcich, Meijerink JP, Van Goudoever JB, et al . Muc2-deficient小鼠自发发展结肠炎,表明MUC2结肠保护至关重要。胃肠病学。2006;131 (1):117 - 29。gydF4y2Ba
杨Velcich, W,嘿,J, Fragale,尼古拉斯·C Viani年代,et al。小鼠的结肠直肠癌遗传缺陷的粘蛋白Muc2。科学(纽约)。2002,295 (5560):1726 - 9。gydF4y2Ba
陆P, Burger-van Paassen N, van der Sluis M, Witte-Bouma J, Kerckaert JP, van Goudoever JB, et al .结肠基因表达模式的粘蛋白Muc2基因敲除小鼠结肠炎中揭示不同阶段发展。Inflamm肠道说。2011;17 (10):2047 - 57。gydF4y2Ba
de Vos WM。微生物简介:Akkermansia muciniphila:守恒的肠道共生有机体充当看门人的粘膜。微生物学。2017;163 (5):646 - 8。gydF4y2Ba
茨C, de Vos WM。微生物内部,从多样性函数:Akkermansia的情况。ISME日报。2012;6 (8):1449 - 58。gydF4y2Ba
Derrien M,沃恩EE, Plugge厘米,de Vos WM。sp. Akkermansia muciniphila将军11月,11月,人类肠道mucin-degrading细菌。Int J系统另一个星球Microbiol。2004; 54 (Pt 5): 1469 - 76。gydF4y2Ba
Ottman N, Reunanen J, Meijerink M, Pietila TE, Kainulainen V, Klievink J, et al . Pili-like蛋白质的Akkermansia muciniphila调节宿主免疫反应和肠道屏障功能。《公共科学图书馆•综合》。2017;12 (3):e0173004。gydF4y2Ba
Plovier H,埃弗拉德,Druart C, Depommier C, Van Hul M,吉尔茨L, et al .纯化膜蛋白从Akkermansia muciniphila或巴氏杆菌改善肥胖和糖尿病老鼠的新陈代谢。Nat医学。2017;23(1):107 - 13所示。gydF4y2Ba
Cani PD,德Vos WM。下一代有益微生物:Akkermansia muciniphila。Microbiol前面。2017;8:1765。gydF4y2Ba
刀MC,埃弗拉德Aron-Wisnewsky J, Sokolovska N, Prifti E,教堂司事EO, et al . Akkermansia muciniphila和改善代谢健康膳食干预中肥胖:与肠道微生物丰富度与生态的关系。肠道。2016;65 (3):426 - 36。gydF4y2Ba
Schneeberger M,埃弗拉德,Gomez-Valades AG)、马塔莫罗斯年代,拉米雷斯,Delzenne NM, et al . Akkermansia muciniphila反向与炎症的发生,改变脂肪组织代谢及代谢紊乱在肥胖老鼠。Sci众议员2015;5:16643。gydF4y2Ba
埃弗拉德一,吉尔茨茨C、L, Ouwerkerk JP, Druart C, Bindels磅,et al。相声之间Akkermansia muciniphila和肠道上皮细胞控制食源性肥胖。《美国国家科学院刊a . 2013; 110 (22): 9066 - 71。gydF4y2Ba
名记van der卢格特曾B, F,琵琶C, Lamprakis,萨拉查E, Boekschoten MV,等。综合分析肠道微生物群组成,主机结肠基因表达和管腔内的代谢物在C57BL / 6 j小鼠衰老。衰老。2018;10 (5):930 - 50。gydF4y2Ba
Langille MG他不CJ, Koenig我Dhanani,玫瑰RA, Howlett SE, et al。衰老小鼠肠道微生物的变化。微生物。2014;2 (1):50。gydF4y2Ba
教唆E, Nylund L, M烛光,Ostan R, Bucci L,扎E,等。通过老化,和超越:肠道微生物群在老年人和百岁老人和炎症状态。《公共科学图书馆•综合》。2010;5 (5):e10667。gydF4y2Ba
教唆E, Franceschi C, Rampelli年代,Severgnini M, Ostan R, Turroni年代,et al .肠道微生物群和极端的长寿。当代生物学:CB。2016;26 (11):1480 - 5。gydF4y2Ba
凡发现AA, Hugenholtz F,梅耶尔B, B最高统治者,Perdijk O, Vermeij WP, et al .前线科学:色氨酸限制逮捕B细胞发展和增强了微生物多样性WT和过早衰老Ercc1 (−/ Delta7)老鼠。J Leukoc杂志。2017;101(4):811 - 21所示。gydF4y2Ba
范发现AA,君主B, Hugenholtz F,梅耶尔B, Hoogerland是的,Mihailova V, et al。补充与乳杆菌WCFS1防止减少结肠粘液屏障的加速老化Ercc1 (−/ Delta7)老鼠。Immunol前面。2016;7:408。gydF4y2Ba
Vermeij WP、Hoeijmakers JH Pothof j .基因组完整性在老化:人类的综合症,小鼠模型,和治疗选择。为Toxicol杂志》2016;56:427-45。gydF4y2Ba
恩格斯N, Wienands j .内存控制由B细胞抗原受体。Immunol启2018;283 (1):150 - 60。gydF4y2Ba
Neef, Sanz y未来益生菌在功能性食品和膳食补充剂的科学发展。当前的临床营养与代谢护理。2013;16 (6):679 - 87。gydF4y2Ba
Elderman M,君主B, F Hugenholtz, Graversen K, Huijskes M, Houtsma E, et al。年龄对肠道粘膜厚度的影响,微生物群组成和免疫小鼠的性的关系。《公共科学图书馆•综合》。2017;12 (9):e0184274。gydF4y2Ba
Derrien M,茨C, de Vos WM。Akkermansia muciniphila和它的作用在调节主机功能。活细胞Pathog。2017; 106:171 - 81。gydF4y2Ba
Pelaseyed T, Bergstrom JH, Gustafsson JK Ermund, Birchenough通用、舒特等人,et al .粘液和黏蛋白的杯状细胞和肠上皮细胞提供的第一防线胃肠道免疫系统并与之交互。Immunol启2014;260 (1):8-20。gydF4y2Ba
Tran L, Greenwood-Van MB。肠上皮屏障的与年龄有关的改造。J Gerontol Sci地中海Sci杂志。2013;68 (9):1045 - 56。gydF4y2Ba
Reunanen J, Kainulainen V, Huuskonen L, Ottman N,茨C, Huhtinen H, et al . Akkermansia muciniphila坚持肠上皮细胞,增强上皮细胞层的完整性。:环境Microbiol。2015; 81 (11): 3655 - 62。gydF4y2Ba
Bodogai M O ' connell J, K金,金Y, Moritoh K,陈C, et al。共生的细菌导致胰岛素抵抗衰老通过激活先天B1a细胞。科学转化医学。2018;10 (467)。gydF4y2Ba
Derrien M, Van Baarlen P, Hooiveld G,诺E,穆勒M, de Vos WM。调制的黏膜免疫反应、宽容和扩散的老鼠殖民mucin-degrader Akkermansia muciniphila。Microbiol前面。2011;2:166。gydF4y2Ba
Groschwitz KR,霍根SP。肠道屏障功能:分子调控和疾病发病机理。J过敏Immunol。2009; 124(1): 3 -测验1 - 2。gydF4y2Ba
Zmora N, Zilberman-Schapira G,苏伊士J,铁道部U, Dori-Bachash M, Bashiardes年代,et al。个性化的肠道粘膜殖民抵抗经验独特益生菌与宿主和微生物特性。细胞。2018;174 (6):1388 - 405. - e21。gydF4y2Ba
Gutzeit C, Magri G, a Cerutti肠道IgA生产及其在宿主交互作用。Immunol启2014;260 (1):76 - 85。gydF4y2Ba
Cote NML Tremblay年代,Grenier G, Duclos-Lasnier G,福捷LC, Ilangumaran年代,et al。回肠抗菌肽表达特异表达。免疫和老化:我& a . 2017;十四19。gydF4y2Ba
海恩斯L, Maue AC。衰老T细胞功能的影响。当今Immunol。2009; 21 (4): 414 - 7。gydF4y2Ba
戴维斯LC,泰勒公关Tissue-resident巨噬细胞:当时和现在。免疫学。2015;144 (4):541 - 8。gydF4y2Ba
戴维斯LC、大米厘米,Palmieri EM,泰勒公关,库恩DB, McVicar DW。腹膜tissue-resident巨噬细胞的新陈代谢将参与微生物利用tissue-niche燃料。Nat Commun。2017; 8 (1): 2074。gydF4y2Ba
Ouwerkerk JP Aalvink年代,茨C, De Vos WM。可行的制备和保存Akkermansia muciniphila细胞治疗干预措施。有益的微生物。2017;8 (2):163 - 9。gydF4y2Ba
Dominguez LJ,市场的生物代谢综合征和老化。当前的临床营养与代谢护理。2016;第5 - 11(1):19日。gydF4y2Ba
尼科尔斯GA,硅藻土EJ。代谢综合征相关组件是未来医疗费用独立于心血管事件住院和糖尿病。金属底座Syndr遗传代数Disord。2011; 9(2): 127 - 33所示。gydF4y2Ba
Salonen, Nikkila J, Jalanka-Tuovinen J, Immonen O, Rajilic-Stojanovic M, Kekkonen RA, et al .粪便DNA提取方法的比较分析与系统发育微阵列:有效恢复使用机械细胞裂解细菌和古细菌的DNA。J Microbiol方法。2010;81 (2):127 - 34。gydF4y2Ba
Ramiro-Garcia J,爱马仕GDA、Giatsis C Sipkema D, Zoetendal如Schaap PJ, et al . NG-Tax,高度精确和验证管道进行分析16 s rRNA扩增子从复杂的生物群落(版本1;裁判:2与保留批准,1不批准)。F1000Research。2016 (5):1791。gydF4y2Ba
警棍L, Shetty s R版本1.1.10013微生物分析的工具。2017年。gydF4y2Ba
McMurdie PJ,福尔摩斯s phyloseq: R包可再生的交互式分析微生物人口普查数据和图形。PloS one。2013;8 (4):e61217。gydF4y2Ba
陆君主B, Loonen LM, P, Hugenholtz F,茨C, Stolte呃,et al . IL-22-STAT3通路中发挥着关键作用在维护回肠内稳态的老鼠缺乏分泌粘液屏障。Inflamm肠道说。2015;21 (3):531 - 42。gydF4y2Ba
萨勃拉曼尼亚,Tamayo P,而VK,慕克吉年代,艾伯特提单,吉列,等。基因集富集分析:基于知识的方法解释全基因组表达谱。《美国国家科学院刊S a . 2005; 102 (43): 15545 - 50。gydF4y2Ba
克莱默,绿色J波拉德J Jr, Tugendreich因果分析方法在独创性途径分析。生物信息学(英国牛津大学)。2014;30 (4):523 - 30。gydF4y2Ba
王X,种子b银行PCR引物进行定量基因表达分析。核酸研究》2003;31 (24):e154。gydF4y2Ba
确认gydF4y2Ba
我们要感谢楼Hugenholtz为她帮助在16 s rRNA测序和珍妮詹森对她帮助在微阵列的性能。我们也感谢Jan Hoeijmakers怀孕他的出色工作gydF4y2BaErcc1gydF4y2Ba−/Δ7gydF4y2Ba小鼠模型。伊薇特房龙被公认为她帮助育种和基因分型鼠标军团。gydF4y2Ba
资金gydF4y2Ba
这项工作是由荷兰科学研究组织(斯宾诺莎奖和暹罗重力格兰特024.002.002)和通过NWO研究生计划在食品结构、消化和健康。gydF4y2Ba
可用性的数据和材料gydF4y2Ba
微阵列基因表达数据可用在NCBI综合(GEO)与GSE126730数量。16 s rRNA基因数据中描述这项研究已经存入顺序读取存档(SRA) NCBI (PRJNA525606)。gydF4y2Ba
作者信息gydF4y2Ba
作者和联系gydF4y2Ba
贡献gydF4y2Ba
AB,对WMV, HS和CB设计研究。小儿麻痹症怀孕的gydF4y2BaErcc1gydF4y2Ba−/Δ7gydF4y2Ba小鼠模型和RB负责繁殖gydF4y2BaErcc1gydF4y2Ba−/Δ7gydF4y2Ba老鼠。提单、AB、SA、BM和b进行了实验。提单写的手稿。AB、SA、BS、小儿麻痹,对WMV, HS、WS和CB导致的修订手稿。所有作者阅读和批准最终的手稿。gydF4y2Ba
相应的作者gydF4y2Ba
道德声明gydF4y2Ba
伦理批准gydF4y2Ba
实验研究与实验动物保健原则和荷兰立法和荷兰瓦赫宁根伦理委员会的批准。gydF4y2Ba
同意出版gydF4y2Ba
不适用。gydF4y2Ba
相互竞争的利益gydF4y2Ba
作者宣称没有利益冲突。gydF4y2Ba
出版商的注意gydF4y2Ba
施普林格自然保持中立在发表关于司法主权地图和所属机构。gydF4y2Ba
附加文件gydF4y2Ba
额外的文件1:gydF4y2Ba
基因表达谱的结肠和回肠(微阵列分析)。只有基因,在统计学上显著不同(gydF4y2BapgydF4y2Ba< 0.05)和有褶皱变化(FC) <−1.2或> 1.2包括团体之间。(XLSX 64 kb)gydF4y2Ba
额外的文件2:gydF4y2Ba
基因集富集分析(GSEA)回肠和结肠的结果。只有大大丰富途径(包括罗斯福< 0.2)。(XLSX 25 kb)gydF4y2Ba
额外的文件3:gydF4y2Ba
相对基因表达(A)再生islet-derived 3β(gydF4y2BaReg3bgydF4y2Ba),(B)再生islet-derived 3γ(gydF4y2BaReg3ggydF4y2Ba),(C) Claudin 2 (gydF4y2BaCldn2gydF4y2Ba),(D) Claudin 8 (gydF4y2BaCldn8gydF4y2Ba),(E)连环蛋白(钙粘着蛋白相关蛋白),α3 (gydF4y2BaCtnna3gydF4y2Ba)和(F) ST6 (alpha-N-acetyl-neuraminyl-2 3-beta-galactosyl-1 3) -N-acetylgalactosaminideα2,6-sialyltransferase 6 (gydF4y2BaSt6galnac6gydF4y2Ba在回肠)。(G)的相对表达C-X-C主题趋化因子配体(13gydF4y2BaCxcl13gydF4y2Ba)、B (H)淋巴激酶(gydF4y2Ba黑色gydF4y2Ba集群的区别),(我)4 (gydF4y2BaCd4gydF4y2Ba72),(J)集群的区别(gydF4y2BaCd72gydF4y2Ba),(K) toll样受体7 (gydF4y2BaTlr7gydF4y2Ba),(L) 12 (toll样受体gydF4y2BaTlr12gydF4y2Ba在结肠)。(PDF 266 kb)gydF4y2Ba
额外的文件4:gydF4y2Ba
存活率和身体的重量gydF4y2BaErcc1gydF4y2Ba−/Δ7gydF4y2Ba老鼠。(一)百分比所有小鼠的生存。这些数据包括12 - 13每组有一个额外的5 - 6组小鼠在16周审查。(B)百分比只雄性老鼠的生存(gydF4y2BangydF4y2Ba= 8 - 10 /组)和(C)雌性老鼠(gydF4y2BangydF4y2Ba= 8 - 10 /组)。(D)在克每周测量体重小鼠(gydF4y2BangydF4y2Ba=每组18老鼠),(E)雄性老鼠(n = 8 - 10 /组)和(F)雌性老鼠(n = 8 - 10 /组)。注:5 - 6雌性老鼠的数量是牺牲在16周。(PDF 261 kb)gydF4y2Ba
额外的文件5:gydF4y2Ba
(一)饮食成分表。(B)与引物序列用于qPCR表。表的列表(C)抗体用于流式细胞术。(多克斯21 kb)gydF4y2Ba
权利和权限gydF4y2Ba
开放获取gydF4y2Ba本文是分布式根据创作共用署名4.0国际许可证(gydF4y2Bahttp://creativecommons.org/licenses/by/4.0/gydF4y2Ba),允许无限制的使用、分配和复制在任何媒介,你提供给适当的信贷原始作者(年代)和来源,提供一个链接到Creative Commons许可,并指出如果变化。知识共享公共领域奉献豁免(gydF4y2Bahttp://creativecommons.org/publicdomain/zero/1.0/gydF4y2Ba)适用于数据可用在这篇文章中,除非另有说明。gydF4y2Ba
关于这篇文章gydF4y2Ba

引用这篇文章gydF4y2Ba
van der卢格特曾,B。,凡发现,嗜、Aalvink年代。gydF4y2Baet al。gydF4y2BaAkkermansia muciniphilagydF4y2Ba改善老年结肠粘液厚度下降,减弱免疫激活加速老化gydF4y2BaErcc1gydF4y2Ba−/Δ7gydF4y2Ba老鼠。gydF4y2BaImmun老化gydF4y2Ba16gydF4y2Ba6 (2019)。https://doi.org/10.1186/s12979 - 019 - 0145 - zgydF4y2Ba
收到了gydF4y2Ba:gydF4y2Ba
接受gydF4y2Ba:gydF4y2Ba
发表gydF4y2Ba:gydF4y2Ba
DOIgydF4y2Ba:gydF4y2Bahttps://doi.org/10.1186/s12979 - 019 - 0145 - zgydF4y2Ba
关键字gydF4y2Ba
- 老化gydF4y2Ba
- Akkermansia muciniphilagydF4y2Ba
- 肠道免疫gydF4y2Ba
- 黏液层gydF4y2Ba
- 肠屏障gydF4y2Ba