隔离的抗炎和上皮加强拟杆菌和Parabacteroides种虫害从健康的粪便捐赠


1
人类微生物组研究项目,医学院,00290年芬兰赫尔辛基,赫尔辛基大学
2
转化基因组学研究所、病原微生物部门,旗杆,AZ 86001,美国
*
作者所对应应该解决。
__
这些作者的贡献同样这项工作。
营养物质
2020年,12(4),935;https://doi.org/10.3390/nu12040935
收到:2020年2月28日/修改:2020年3月23日/接受:2020年3月24日/发表:2020年3月27日
(这篇文章属于部分益生元、益生菌)
文摘
:
改变肠道微生物群与系统性和肠道疾病有关,如炎症性肠病(IBD)。Dysbiotic微生物群与增强促炎的能力特点是厌氧的消耗共生体,兼性厌氧细菌的比例增加,以及减少多样性和稳定性。在这项研究中,我们开发了一个体外高通量筛选鉴定分离肠道共生的细菌与抗炎能力健康的粪便微生物群移植供体。刚分离肠道细菌筛选的能力减弱大肠杆菌全身的脂多糖(LPS)白介素8(引发)释放HT-29细胞。产生了一系列的屏幕拟杆菌和Parabacteroides隔离,标识为p . distasonis,b . caccae,b . intestinalis,b .均匀化,b . fragilis,b . vulgatus和b . ovatus使用全基因组测序。我们观察到一个信息与上皮细胞体外炎症没有必要缓解了文化隔离的媒体也是有效的抗炎作用并没有与分离的肠上皮细胞粘附能力。抗炎隔离也对肠上皮细胞单层加强行动,缺乏必要的基因合成hexa-acylated,促炎的脂质,有限合伙人的一部分。然而,抗炎效应分子仍有待确定。的拟杆菌菌株分离和特征在这项研究中有潜在的作为所谓的下一代益生菌。
1。介绍
人类的肠道也有复杂的细菌生态系统的物种的90%属于厚壁菌门和拟杆菌门1,2,3]。大多数人把大约200不同种类的细菌在胃肠道不同丰富但只有大约30%的物种,所谓的共同核心微生物群,之间共享对象(4,5]。与宿主的遗传和免疫背景,环境因素如饮食,使用抗生素和其他药物和卫生条件影响肠道微生物群组成(6]。Dysbiotic、不平衡肠道菌群组成和增强促炎的能力的特点是减少物种丰富度和多样性以及减少微生物稳定性(7]。health-associated变化丰富,短链脂肪酸制造酸性物质厚壁菌门和促炎,脂多糖(LPS)——包含pathobionts gamma-Proteobacteria等与失调(8]。此外,肠道免疫内稳态和屏障功能与系统性和肠道疾病相关,如炎症性肠病(IBD)和肠易激综合症(IBS) (9]。多项研究表明,炎症性肠病病人减少了大量的粘膜或粪便拟杆菌健康受试者相比(10,11,12]。
操纵的肠道微生物群朝着更加平衡和优化微生物群落,促进人类健康近年来已经受到了越来越多的兴趣。除了传统的益生菌和益生元13),新的治疗方法从操纵总微生物群粪便微生物群移植(FMT, [14])窄谱微生物群操纵的靶向抑制代谢途径在一个特定的细菌集团(15正在接受调查。FMT治疗复发性的巨大成功Clostridioides固执的感染(rCDI),并承诺发现几个其他条件(14)一起研究微生物群失调为所谓的下一代益生菌和细菌疗法的发展8]。在炎症性肠病,治疗成功率FMT的变量(16,17),使用细菌社区定义为一种强有力的细菌疗法可以提供更加一致的疗效相比FMT捐助者之间有显著差异(18]。事实上,在炎症性肠病的小鼠模型,用三重治疗拟杆菌应变的组合,特别是b . ovatus,减少炎症和更有效的减少比传统FMT结肠炎(19]。最近,美国食品和药物管理局(FDA)批准了一个试验性新药(印第安纳州)应用程序的安全性和有效性b . ovatus,亚种和b . vulgatus作为治疗rCDI [19]。在一项研究中关于封装FMT,拟杆菌和Parabacteroides的物种,主要导致了供体移植,可能作为关键物种在维护donor-like微生物群(20.]。
拟杆菌门后第二大门壁厚菌门在人类肠道健康的成年1,2,3),其相对比例的总在粘膜微生物群是高于粪便(21]。属拟杆菌,占大约30%的肠道微生物群,包含最主要的物种在细菌性的顺序(22]。拟杆菌种虫害配有不同的机制,以小肠的适应恶劣的环境,如代谢许多饮食host-derived多糖,氧气使用双相障碍细胞色素氧化酶宽容,和大量表达的细胞表面结构(23]。拟杆菌物种包括很多重要的机会致病菌,但基本平衡的微生物群的成员,他们被认为是health-maintaining。这是由于他们的能力加强的上皮屏障,改善炎症产生抗炎分子如多糖(PSA),鞘脂类和外膜囊泡(OMV)提到的分子的运输通过厚厚的粘液上皮细胞层(24,25,26,27]。因此,拟杆菌种虫害可能承诺bacteriotherapeutic候选人在临床使用和营养的应用程序,但需要更多的研究在这个问题上。安全评估的压力需要彻底因为一些属的代表机会致病菌也很重要。
在这项研究中,我们旨在分离肠道共生的细菌与抗炎能力从健康FMT捐助,建立体外高通量筛选试验。在筛选中,我们评估了潜在的新孤立的肠道细菌减弱LPS-induced白介素8(引发)从肠上皮细胞即释放。,以缓解炎症反应引起的有限合伙人。有前途的隔离从屏幕上被净化,彻底的验证其抗炎能力后,他们被部分初步确认16 s rRNA基因测序。隔离代表拟杆菌和Parabacteroides种虫害被选作进一步的研究,包括全基因组测序和评估的胶粘剂和上皮细胞强化属性。
2。材料和方法
2.1。细菌分离和初步鉴定
筛选出来的粪便从一个健康的成年人,充当了捐赠者的FMT,冻存作为一个盐水- 10%甘油溶液在−80°C使用协议成功地用于粪便银行FMT [28]。准备做下环境空气和示例被冻结在1.5 h排便。使用捐赠者的经医院伦理委员会批准的芬兰赫尔辛基和有所(DnroHUS124/13/03/01/11)。一项研究[描述的捐赠,21),提供一份书面知情同意。捐献者是一个健康的芬兰女,42岁,有一个正常的身体质量指数(BMI < 25),没有胃肠道症状和抗生素的使用在过去的6个月。捐赠者通过粪便捐助者的所有筛选试验(28]。所有的实验都是按照指导方针和有关规定进行。冷冻的粪便的解决方案是解冻在4 - 5小时,连续稀释在磷酸盐(PBS)和培养布鲁氏菌琼脂(西格玛奥德里奇,圣路易斯,密苏里州,美国)补充5%的羊血(生物Karjalohja男孩,Karjalohja,芬兰),岐阜厌氧介质(GAM;Nissui制药有限公司,日本东京)和钢筋梭菌的培养基(RCM;美国,正欲火花)在厌氧条件下使用一个厌氧室含85% N2,10%的公司2,5%的H2在37°C(概念+厌氧工作站,Ruskinn科技有限公司,布,英国)。内的厌氧室自动清除气体室。钯催化剂在室回收任何残余O2和厌氧颜色指示条被用来验证厌氧条件。所有媒体都减少了厌氧大气中的厌氧工作站前24小时内种植。独立的殖民地了超过72小时和re-streaked新的琼脂板上。每个隔离在适当的肉汤或琼脂培养,即。,the medium used originally to isolate the bacterium, for the preliminary screening of attenuation capacity. Frozen stocks were prepared from the same bacterial culture and stored at −80 °C. The screening of anti-inflammatory isolates (described in detail in paragraph 2.3) was first carried out in 96-well microtiter plates using one replicate due to the large number of isolates. Next, the screening was repeated for potentially positive isolates using three replicates. The potentially positive isolates were re-streaked on agar plates until pure cultures were obtained. The pure cultures were subjected to Gram-staining and microscopy to evaluate the purity of the cultures. Partial 16S rRNA gene sequencing was used for tentative identification, for which bacterial mass from each isolate was resuspended in TE buffer (10 mM Tris-HCl, 1 mM EDTA, pH 8) and heated to 95 °C for 15 min to break the cells. The resulting cell lysates were used as the template in PCR amplification of the partial 16S rRNA gene using 27F DegL (5′-AGR GTT YGA TYM TGG CTC AG-3′) forward and Pd (5′-GTA TTA CCG CGG CTG CTG-3′) reverse primers. Sanger sequencing of the PCR product using the forward primer was carried out in the Institute of Biotechnology core facility, University of Helsinki. The partial 16S rRNA gene sequences were compared to NCBI 16S ribosomal RNA sequences database using BLASTn to acquire tentative identification at the genus level.
2.2。上皮细胞系
人类结肠上皮细胞系Caco-2 169 (ACC)和HT-29 299 (ACC)从德国获得微生物和细胞培养的集合(DSMZ)和生长在孵化器37°C下好氧的气氛有限公司为5%2。细胞达到80%后通过融合使用HyClone HyQTase(美国通用电气医疗集团生命科学、马尔堡)分离细胞和通道数字28被用于实验。Caco-2细胞培养RPMI 1640中(美国Sigma-Aldrich)补充heat-inactivated(30分钟56°C)胎牛血清(20%;的边后卫;英国Gibco),不必要的氨基酸(1%,NEAA;Lonza, Bornem,比利时),消息灵通的15毫米(Lonza、比利时),100 U mL−1青霉素和链霉素(害虫;Lonza、比利时)和2毫米谷酰胺(Lonza、比利时)。5 HT-29细胞生长在本人(Lonza、比利时)中含有10%的边后卫和100 U mL−1害虫。
2.3。筛选分离株的抗炎能力
细菌分离株的能力减弱LPS-induced引发生产HT-29细胞测量如前所述[29日),但分析被调整至高吞吐量96孔板格式。HT-29每口井的细胞被播种10000到96孔酶标衰减试验。肉汤培养的细菌都洗一次本人5中补充的边后卫(10%)和resuspended在同一介质。分离的细菌大量生长在固体琼脂收集和resuspended McCoy 5一个中等的边后卫补充。OD细菌悬浮液进行调整600海里0.25对应的约108细胞/毫升和100µl暂停添加到8天post-plating HT-29细胞和孵化在37°C下1小时含氧的气氛有限公司为5%2。实验中使用的细菌浓度(最终的浓度107细胞/毫升)在体外实验中被认为是生物相关的数量拟杆菌种虫害的肠理论上可以5×109/ 3×10毫升10/毫升。大量的拟杆菌门在一个健康的微生物群有很多inter-individual方差,但总的来说它是30%在粪便和粪便中的细菌含量大约是1011/ g的粪便30.]。的百分比拟杆菌种虫害的粪便捐赠用于本研究大约5% (21]。本人5中含有10%的边后卫是用作控制。之后,细菌悬浮液被删除从HT-29单层和200年µl McCoy 5 1 ng / mL的媒介大肠杆菌有限合伙人(西格玛奥德里奇,美国)补充道。与有限合伙人,4小时孵化后上层清液收集和引发的水平是衡量使用ELISA测定(美国圣地亚哥BD OptEIATM组)。有氧条件下增加公司的使用2(5%),筛选试验使用适当厌氧细菌被认为是由于激活时间短。之前,小时孵化HT-29细胞移除细菌细胞和添加有限合伙人。也知道,厌氧细菌可以容忍氧1小时到72小时(根据物种31日),而拟杆菌种虫害维持有氧条件比其他厌氧肠道细菌(32]。
引发水平计算使用四个参数逻辑曲线和隔离的效果在LPS-induced引发生产评估通过比较水平的有限合伙人控制。在第一个筛选每个孤立使用只有一个复制()和积极的隔离标准减少引发水平低于有限合伙人控制。没有统计测试应用每个隔离测试只有一个复制。在第二轮的筛选,三个复制(井)隔离使用,平均每个样本三个复制的结果是有限合伙人相比控制通过t检验,和所有隔离显示显著(p< 0.05)减少IL-8-levels相比控制选择进行进一步的测试。无二次统计分析使用因果测试假阳性拍摄,因为所有的隔离会随后进行验证通过使用生物测试,因此宽松的统计标准应用在筛选阶段。最后,衰减检测与净化,重复多次潜在积极隔离。引发试验只是使用HT-29细胞系由于执行缺陷Caco-2细胞系的TLR4信号造成反应迟钝LPS刺激(33,34,35]。
在介质衰减试验进行了同样如上所述。积极抗炎action-exerting隔离都生长在GAM肉汤在厌氧条件下和OD600海里调整到1.0。调整细菌悬浮液离心(10000 RPM, 3分钟。)颗粒细胞和上层清液与0.2µm过滤器过滤,删除任何剩余的细菌细胞。上层清液稀释至1:4,1:2使用本人5中补充的边后卫(10%)。GAM和本人5被包括在试验控制。
2.4。细菌粘附细胞系和粘液
细菌分离株的粘附Caco-2和HT-29细胞株post-plating(8天)和粘液研究如前所述[36,37]。四个技术复制(平行井)被用于每个实验。细菌生长在一个适当的媒介补充μL 10毫升−1(6′36 H]胸苷(17日Ci更易−1美国珀金埃尔默,沃尔瑟姆)新陈代谢radiolabel细胞。同一物种内的所有隔离种植相同的附着力和测试相同的实验。上皮细胞粘附试验,HT-29 Caco-2细胞被播种10000细胞/ 96孔酶标上。评估坚持肠道粘液从细胞系(单独分析),猪粘液(Sigma-Aldrich 50μg以上在PBS)获准Maxisorp吸附微量滴定板井一夜之间在4°C。[3 h] Thymidine-labelled细菌细胞与适当的清洗介质(本人为HT-29 5细胞,RPMI 1640 Caco-2细胞和粘液化验PBS) OD没有补充和调整600海里0.25,大约108细胞/毫升。细菌悬浮液孵化一小时在上皮细胞单层或粘液在37°C有限公司2孵化器,紧随其后的是洗涤三次删除non-adherent细胞。粘细菌细胞溶解1% sds - 0.1 M氢氧化钠和放射性测量液体闪烁体(Wallac Winspectral 1414年,珀金埃尔默,沃尔瑟姆,妈,美国)。绑定细菌的比例计算相对于细菌悬液的放射性最初添加到井中。
2.5。细菌对上皮的完整性
Caco-2细胞接受enterocytic分化和表达细胞间交叉的复合物,使他们一个合适的模型来衡量transepithelial电阻(TER)作为单层完整性的指标(38,39,40]。细菌分离株的影响后的Caco-2研究了单层如前所述[29日,37]。短暂,50000年Caco-2细胞被播种在宠物插入的孔隙大小3嗯(Sarstedt、Numbrecht、德国)和增长了8天。媒介在每3天了。8天后Caco-2细胞形成单层,部分有区别,在一个适当的增长阶段测量transepithelial耐药性的发展,以及它是如何受外部因素影响(36,37]。大肠杆菌全球已被证明影响单层完整性和用作负控制实验(37]。细菌细胞与RPMI介质和100年洗一次μL OD的细菌悬液调整600年0.25添加到时间后插入0测量。TER测量进行了使用一个EVOM上皮voltohmmeter与电极(英国世界精密仪器)。包含细菌分离株的插入在37°C孵化有限公司2孵化器和TER 24小时后再测量。空白的阻力(测量时间点(0)减去从孵化24小时后进行的测量,和单位面积电阻(Ω厘米2)是由组织的阻力值乘以计算过滤膜的表面积。超过24小时后的变化是通过比较计算样品中控制。
2.6。基因组测序
基因组DNA提取细菌细胞颗粒使用DNeasy血液和组织工具包(试剂盒、希尔登,德国)制造商的协议后,革兰氏阴性细菌。DNA是分散使用Q800R2超声发生器(美国QSonica新城,CT)约500个基点,和基因组库准备使用NEBNext双末端测序®超™II工具包(美国新英格兰生物学实验室,伊普斯维奇,MA)和量化使用图书馆量化工具(KAPA生物系统,沃尔瑟姆,妈,美国)。图书馆汇集在MiSeq克分子数相等的金额和测序(Illumina公司,Inc .,圣地亚哥,美国)。读取数据被存入NCBI SRA下BioProject PRJNA575760。短的读取数据组装使用无人驾驶的基因组组装管道(UGAP;https://github.com/jasonsahl/UGAP),它使用黑桃基因组汇编。每个基因组的物种被发现使用巨妖(PMID: 24580807),证实了其在系统发育树的位置为每个物种生成(见下文)。
全基因组单核苷酸多态性(SNP)输入(WGST)是用来确定任何基因组从这项研究有很高的身份与以前公布的基因组。WGST, SNP矩阵来识别点突变之间的隔离(因此推断毒株亲缘)生成与NASP [41),读取对齐到一个公开可用的组装使用BWA [42]。单核苷酸多态性被称为基因组分析工具包(GATK) [43)和只包含在进一步的分析如果他们出现在所有样本,b)由≥10倍深度≥90%的共识在每个示例中,c)并不是在任何确定的参考基因组复制地区NUCmer [44]。由此产生的SNP矩阵组成的核心基因组共同所有样品分析。最大似然与IQ-TREE系统发育分析45]。最大的吝啬与大型分析v7.0 (46)或RAxML(8.2.10版本,(47])。
2.7。统计分析
所有的附着力和衰减实验使用三到四个技术复制根据试验和重复两到四次(生物复制)来确认结果。细菌的不同文化和不同段落的细胞系中使用了独立的实验。两个示例t检验是用于确定样本之间的显著差异和控制。方差齐性测试执行与列文的测试确定平等或不平等的方差。点二列相关计算衰减之间的二分变量和粘附能力和连续变量。所有统计分析进行了使用IBM SPSS统计程序21.0版(IBM公司,纽约,纽约,美国)的p被认为具有统计显著性的值< 0.05。
2.8。数据可用性
全基因组序列读取数据被存入NCBI SRA下BioProject PRJNA575760。
3所示。结果
3.1。高通量筛选方法
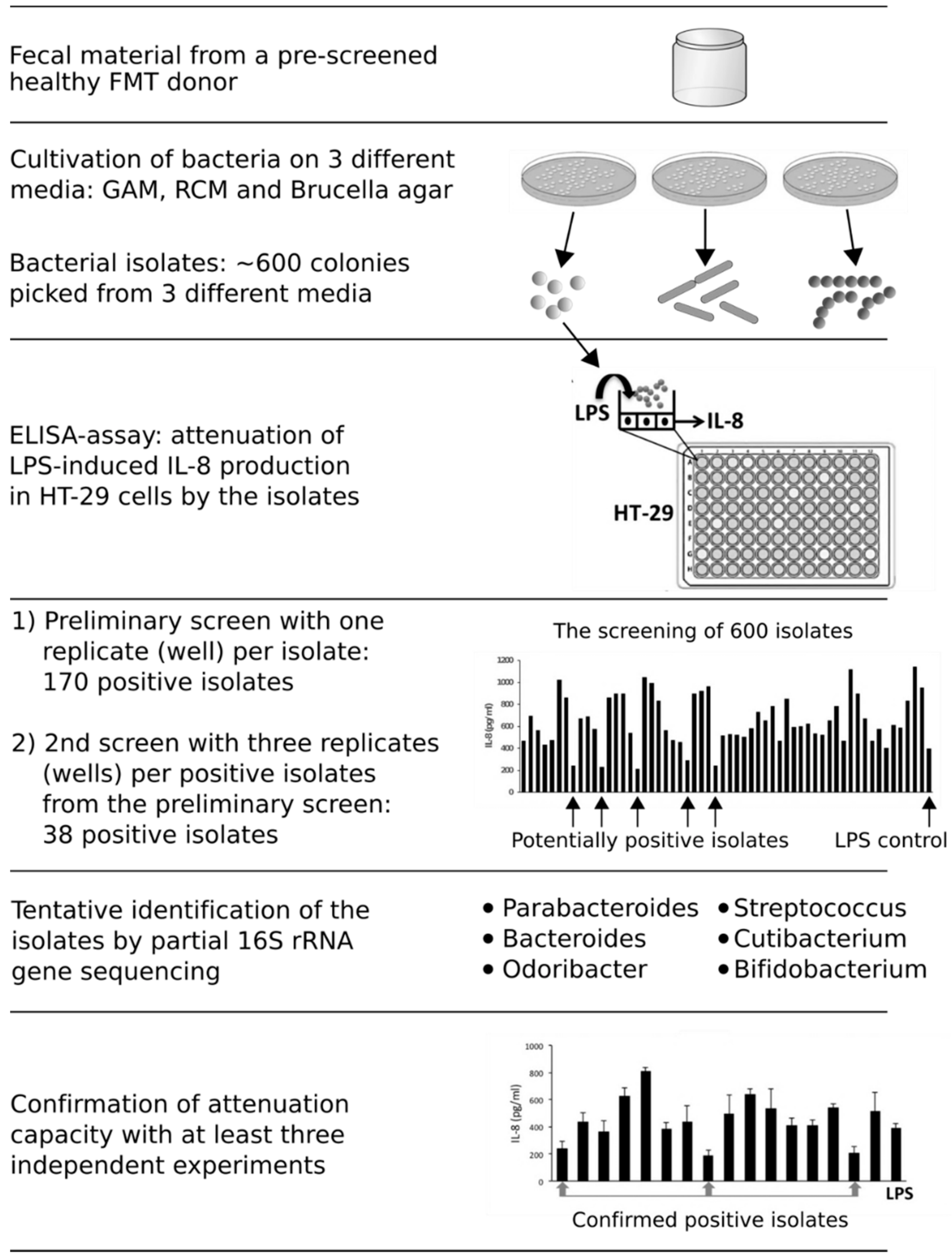
在这项研究中,我们开发了一种高通量分析筛选共生的细菌在人类胃肠道中发挥抗炎作用。首先,筛选出来的细菌的排泄物,健康FMT供体培育在GAM(岐阜厌氧培养基),RCM(钢筋梭菌的培养基)和布鲁氏菌在厌氧条件下琼脂和600细菌隔离了72小时的孵化期间(图1)。筛选抗炎能力首先是由预处理HT-29细胞系的细菌分离,然后用大肠杆菌有限合伙人,导致一个引发释放肠上皮细胞的细胞系。在LPS-induced HT-29细胞是预处理的一个孤立的细菌没有预处理(即比控制。单独LPS-induction),减少产生引发水平被认为是抗炎能力的象征。在第一轮的筛选,我们测试了600年与一个复制()/隔离,隔离170隔离(隔离)的28%减少了引发相比有限合伙人控制水平。第二个屏幕使用几个复制/隔离了38个积极的隔离,大大减少了引发释放HT-29细胞。积极的分离纯化和初步确认在属级使用16 s rRNA基因测序。超过75%的隔离属于细菌性的秩序,包括属Parabacteroides(6)隔离开来,拟杆菌(22隔离)Odoribacter(1)隔离。剩下的隔离被确定为Cutibacterium(3)隔离开来,链球菌(1)隔离双歧杆菌属(5隔离)。我们集中我们的进一步研究Parabacteroides和拟杆菌隔离,形成一个主要细菌分离株的一部分,特别是与FMT后恢复粘膜微生物群(20.,21]。此外,这些物种在先前的研究显示有趣的免疫调节能力12,19,48,49,50,51]。
3.2。全基因组测序(WGS)
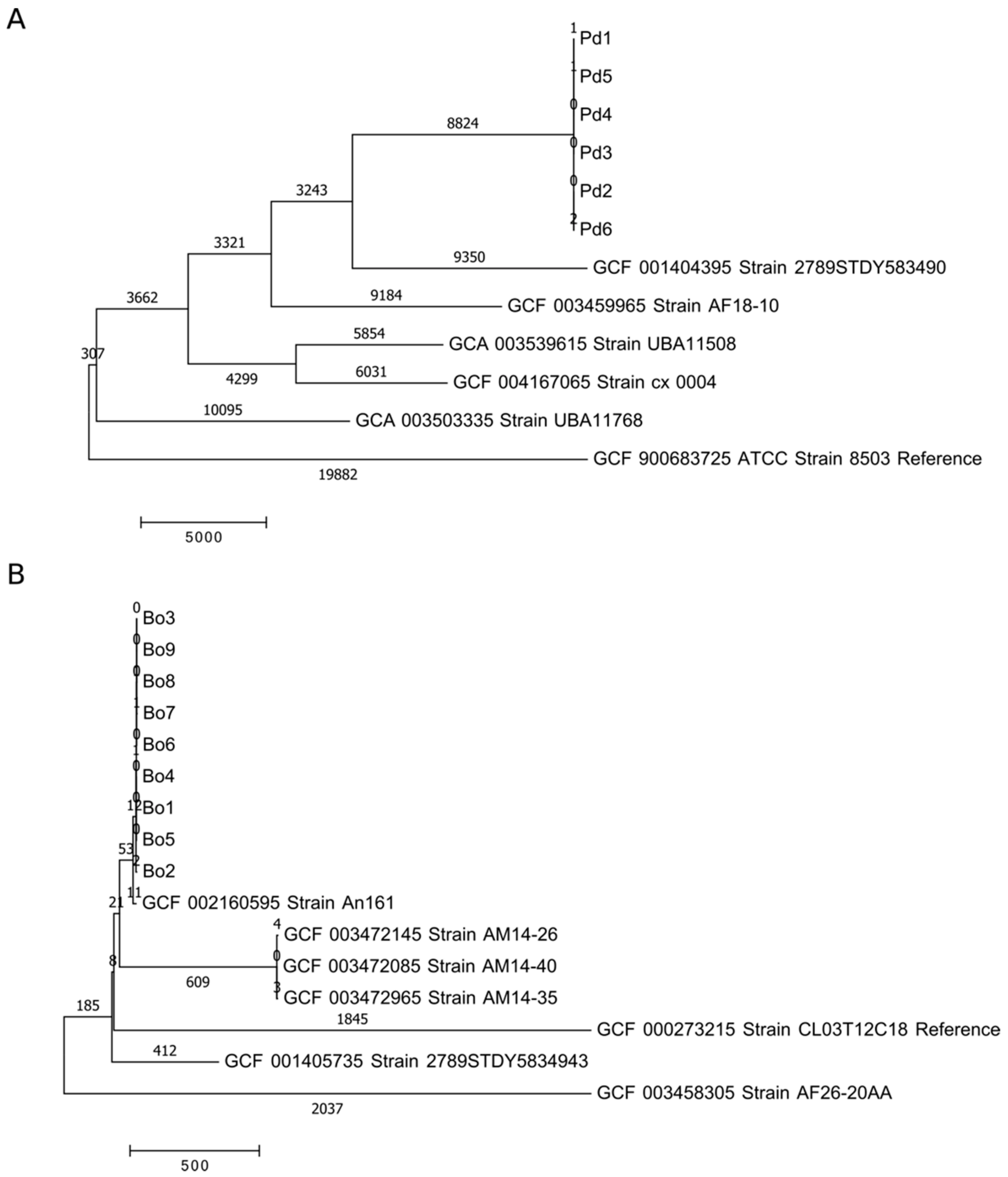
28岁的基因组Parabacteroides和拟杆菌分离得到使用全基因组测序(WGS)和确定一个物种与巨妖(PMID: 24580807;补充表S1)。所有六个Parabacteroides隔离有5.0 Mb组装基因组大小和GC含量和被证实为45.1%Parabacteroides distasonis(Pd)。的拟杆菌隔离被确定为b . caccae(公元前3隔离),b . fragilis(Bf),b . intestinalis(Bi、3隔离)b .均匀化(部),b . vulgatus(Bv 5隔离)b . ovatus(Bo, 9隔离)组装基因组大小和GC含量4.4 - -6.9 Mb和41.9 - -46.5%之间不等。此外,吝啬树基于全基因组单核苷酸多态性(SNP)输入(WGST)表示一个隔离在一个物种的亲缘密切(图2和补充图S1)。此外,SNP打字使用NASP透露,该菌属于同一物种还不到10个基因SNP彼此的差异,但差异超过10000个SNP数据库菌株(平均补充Excel文件,图2和图S1)。因此,所有隔离在同一物种很可能代表相同的应变或至少有一个共同的起源。
3.3。抗炎作用
接下来,我们评估的健壮性免疫调节属性中28隔离,这是他们能够持续缓解LPS-induced引发释放到另一个从一个实验。实验重复三次以上所有的纯化和基因组测序的细菌隔离。明显的隔离(p≤0.05)减少引发释放在至少三个不同的实验,在大多数独立实验(例如,在三个五个实验)被认为是衰减能力(表1)。所有b . caccae,b . intestinalis均匀化和b . vulgatus隔离一再显示,能够显著减少LPS-induced引发释放HT-29细胞相比,有限合伙人控制。在b . caccae,b . intestinalis和b . vulgatus隔离,衰减能力似乎非常稳定和一致的同一物种的隔离。的b . fragilis无法验证隔离发挥抗炎作用,并不影响引发生产LPS-induced HT-29细胞。有趣的是,只有三个六p . distasonis和两个b . ovatus隔离被证实对体外抗炎作用,这表明衰减能力不是一个健壮的特性在这些分离株中,来源于共同的祖先应变和似乎容易失去或易变地表达特征。重要的是,等幅隔离,无显著(p≥0.05)影响有限合伙人控制注意到,没有刺激炎性反应在HT-29细胞系。
3.4。粘膜粘附的隔离
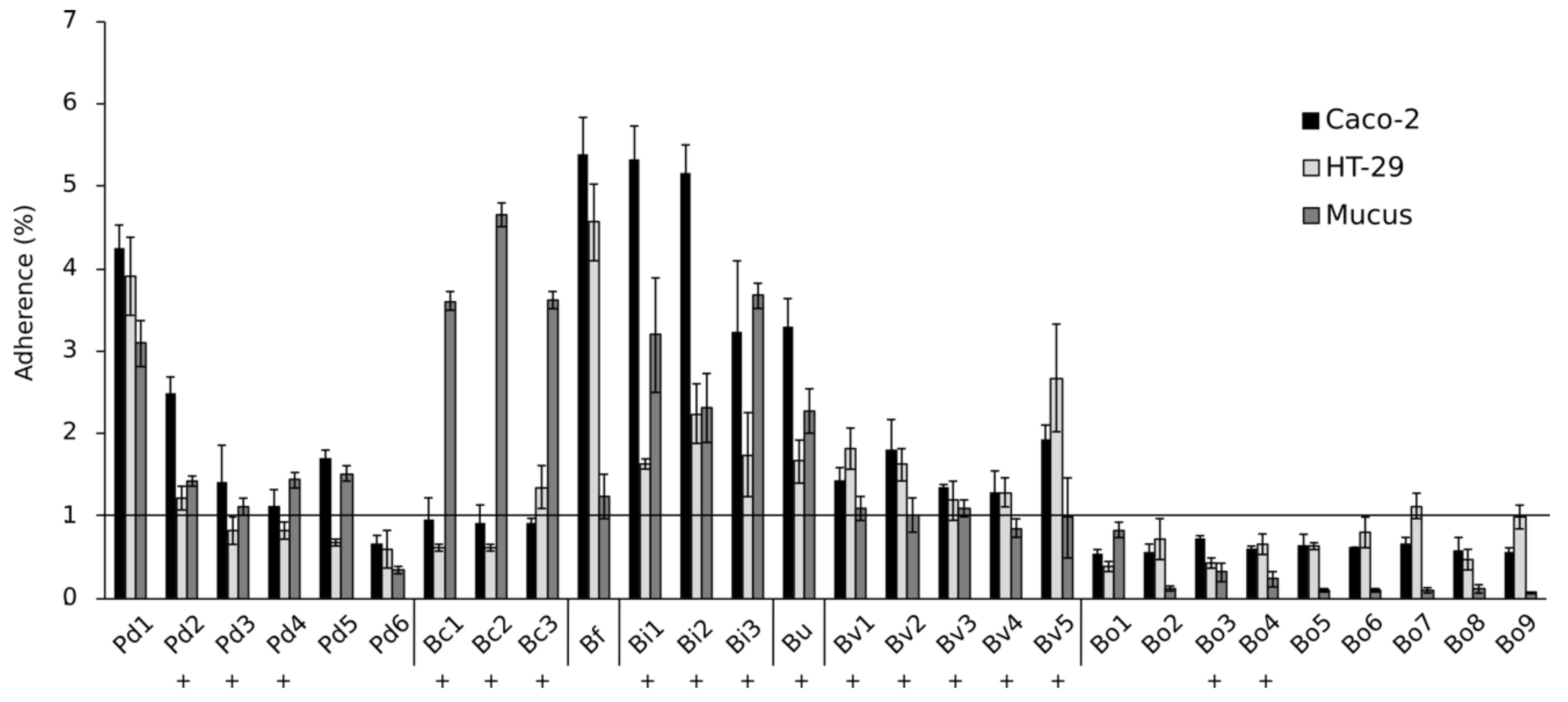
所有b . ovatus分离很明显non-adherent Caco-2 HT-29细胞系以及粘液,因为他们相对坚持比例(衡量粘细菌的比例相对于总添加细菌)低于1%,可以认为是不具体的绑定与背景水平(图3)。这三个b . caccae隔离坚持粘液。在p . distasonis压力,只有Pd1附着而绑定的其他能力p . distasonis隔离接近背景水平。相反,所有b . intestinalis,b .均匀化和b . fragilis隔离可以坚持Caco-2 HT-29和粘液相对坚持约2 - 5%的水平。我们没有发现相关性隔离的能力遵守HT-29细胞和减弱LPS-induced引发释放肠上皮细胞细胞系(biseral相关,rpb= 0.084,p= 0.670),因此抗炎作用似乎并不需要一个公司信息附件细菌和肠上皮细胞之间。
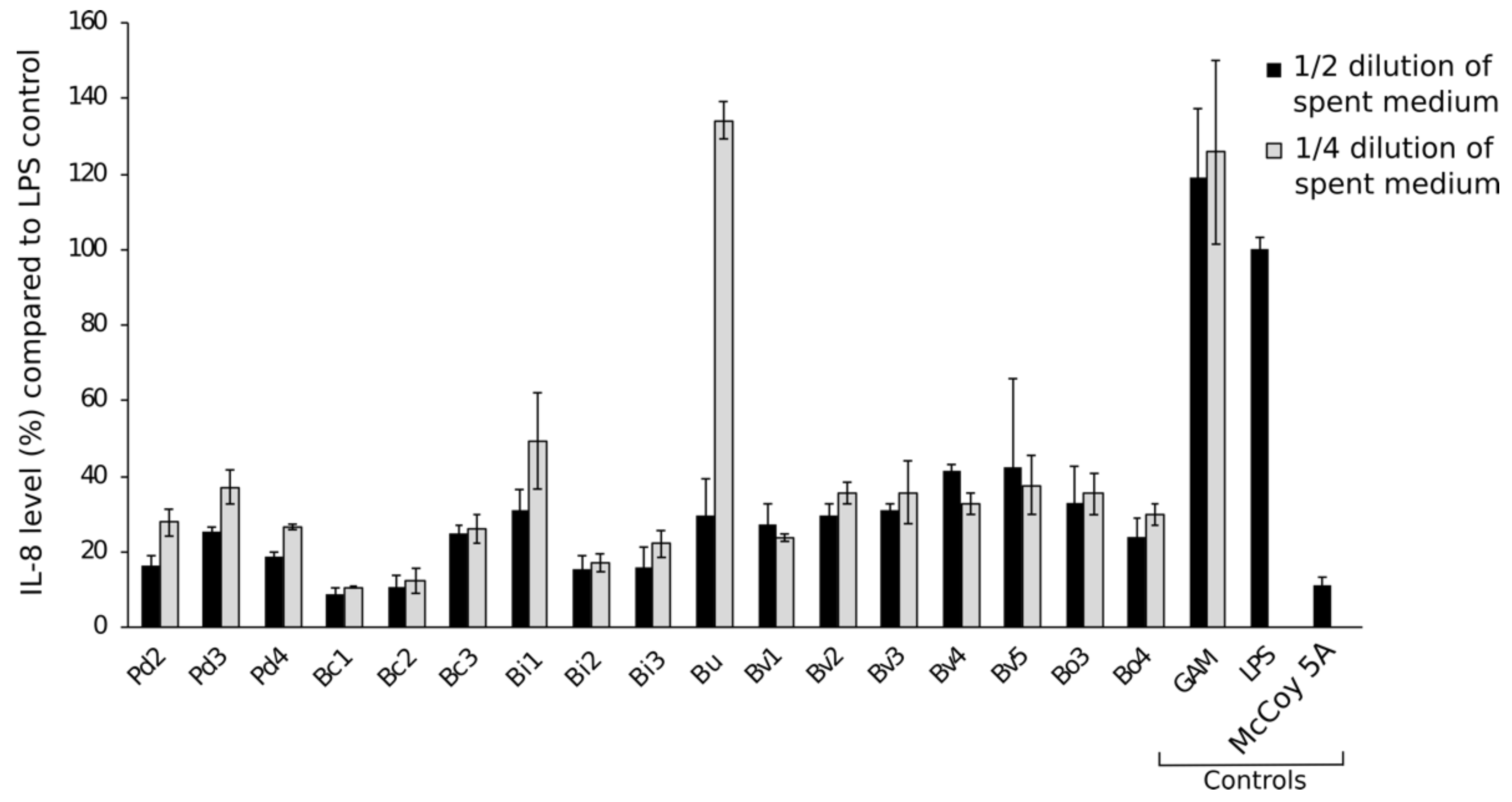
随后,我们评估了游离的抗炎作用的媒介使用文化上层清液细菌培养的衰减试验(图4)。Pre-incubation HT-29细胞与1:2(50%)或1:4(25%)稀释了培养基的分离显著减少引发释放在随后LPS刺激相比,有限合伙人和没有用完的细菌生长介质控制,因此确认公司信息附件细菌和上皮细胞之间没有必要发挥抗炎作用。
3.5。上皮细胞强化行动
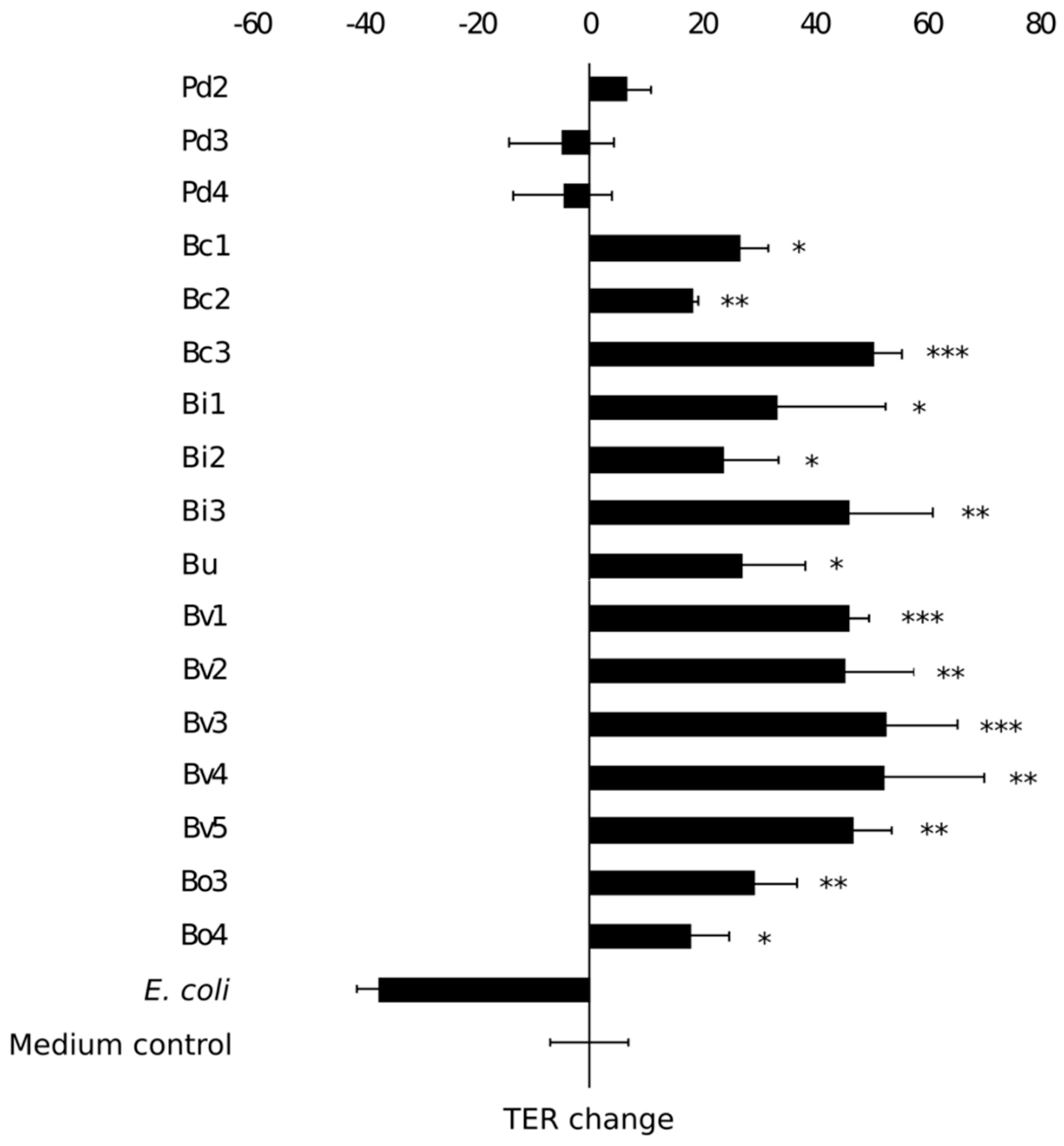
接下来,我们研究了上皮细胞分离的强化属性显示抗炎特性。隔离的能力提高Caco-2单层完整性评估通过培养肠上皮细胞细胞的细菌分离和使用transepithelial电阻测量解决维护或改善屏障功能(TER,图5)。一个商业大肠杆菌实验室菌株是用作负控制不利影响单层的完整性(37]。经过八天的post-plating Caco-2单层显示一个基线TER 319±16Ω。大多数隔离,显示能力减弱LPS-induced引发释放还显示能力增强上皮完整性体外(图5)。作为一个例外,三人p . distasonis隔离并没有引起重大变化值比单层后只接受媒介。换句话说,隔离并没有加强也妥协了单层上皮的完整性。b . vulgatus最有效的分离提高了单层的完整性。在隔离中,能力减弱LPS-induced引发积极与生产相关的能力加强的完整性Caco-2单层(rpb= 0.397,p= 0.061)。然而,之间没有相关性的依从性水平隔离Caco-2细胞和他们的能力加强肠上皮细胞单层完整性(r = 0.109,p= 0.620)。
3.6。拟杆菌的基因在宿主的相互作用。
我们筛选基因组组装数据分离的基因已知宿主相互作用的一个重要的角色拟杆菌spp。首先,我们搜索的基因组基因编码酶的分离Kdo2-lipid修改使用爆炸(基本的局部比对搜索工具,blastp算法)。所有的基因组Parabacteroides和拟杆菌分离株进行基因编码LpxA-LpxK Kdo2脂质合成一部分,但缺乏LpxL基因和LpxM酰基转移酶。LpxL和LpxM需要添加二级酰基链(hexa-acylation)脂质产生一个高度促炎典型gamma-Proteobacteria包括有限合伙人结构大肠杆菌(52,53]。无法hexa-acylate脂质可能解释了为什么Parabacteroides和拟杆菌隔离,尽管LPS-carrying生物,没有引起肠上皮细胞的炎性反应。
当我们发现信息的衰减能力独立接触和可能通过分泌效应分子如两性离子介导的荚膜多糖(保证,如多糖b . fragilis在omv)运输,我们搜查了基因组wcfR同系物的基因编码的蛋白质在ATTGal-ZPS蛋白质的合成是必要的。一个匹配wcfR基因的基因组中发现了b . fragilis隔离(90.9%核苷酸身份,数据未显示),但没有其他隔离,因此不能解释他们的抗炎作用。
我们也搜索的基因组Parabacteroides和拟杆菌分离株的基因编码丝氨酸palmitoyltransferase (SPT BT_0870),一种酶调节鞘脂类的合成,最近报道调解拟杆菌宿主相互作用在维持体内平衡和肠道共生24]。一个匹配的87%、88%和76%核苷酸的身份spt基因组的基因被发现b . caccae,b . ovatus和b . intestinalis分别隔离。73%的匹配spt基因组中还发现的b . fragilis。没有观察到显著的相似性p . distasonis,b .均匀化或b . vulgatus基因组有关spt基因。因此,的存在与否spt基因不解释抗炎或上皮加固隔离的能力。总而言之,我们无法确认所有孤立的存在wcfR或spt,以前与消炎的拟杆菌spp。因此,这些基因似乎并没有普遍分布在拟杆菌种虫害、属性编码可以怀疑调解在一些但不是所有菌株消炎。
4所示。讨论
在这项研究中,我们开发了一种高通量方法筛选肠道共生的潜在抗炎和检索在纯文化38隔离,大部分属的代表拟杆菌和Parabacteroides。我们承认筛查的研究仅限于那些细菌生长在文化条件下应用,但有多个物种出现在人类肠道微生物群,可以发挥抗炎作用,也包括潜在新物种(8]。拟杆菌密切相关的属高度丰富的共生体在人类肠道及其丰度下降与肠道炎症如在炎症性肠病和囊性纤维化患者11,12,54,55),及其丰度高,特别是b . fragilis和b . finegoldii,与一批捐赠凳子上有效地保持在缓解溃疡性结肠炎(UC) FMT [56]。此外,一些老鼠研究最近展示的潜力拟杆菌种虫害在改善肠道炎症19,48,51,57,58]。因此,潜在的拟杆菌种虫害和细菌密切相关,以缓解炎症体内体外似乎对应于他们的行动。
基于WGS结果,我们从FMT捐赠被确定为屏蔽隔离p . distasonis,b . caccae,b . fragilis,b . intestinalis,b .均匀化,b . vulgatus和b . ovatus这些都是典型的在人类肠道肠道共生的。SNP输入显示隔离属于同一物种的代表相同的应变或至少有一个共同的祖先。我们的研究结果和结论符合先前的观察,单一菌株通常占据大多数物种在肠道微生物群(59]。有关拟杆菌spp。即使在多个菌株在一个物种共存的情况下,主应变的相对丰度为87%和91%b .均匀化和b . vulgatus分别为(59]。值得注意的是,拟杆菌和Parabacteroides物种基因非常符合平均只有0.45%的核苷酸变异菌株之间,这可能会阻碍应变分化。另一方面,随着越来越多的基因塑料等肠道共生的普氏菌,差异达到2.44%59]。然而,我们彼此隔离还不到10个SNP差异在同一物种,因此我们认为他们有一个共同的祖先或表示相同的压力。
体外能力减弱炎症被证明物种中分离的菌株中是一致的b . caccae,b . instestinalis和b . vulgatus因为所有的隔离水平显著降低了细胞因子引发LPS-induced肠上皮细胞细胞系。在的情况下p . distasonis和b . ovatus分离株之间的衰减能力不同,尽管它们在基因上非常相似,和详细原因仍有待研究。尽管在栽培条件的一致性,我们不能排除在不同批次培养基因表达的变化。另一个可能性是一些孤立的失去了能力所需的效应分子,仍有待研究和更详细的比较基因组的方法。
3的6p . distasonis隔离能够减弱炎症在我们的体外模型,尽管他们没有提高Caco-2单层上皮的完整性。一般来说,在肠道共生的高度毒株特异性抗炎属性,但是p . distasonis菌株与前面描述的消炎作用。最近的一项研究表明,膜的一部分p . distasonis引起了引发生产减少55%和29%大肠杆菌分别LPS诱导HT-29和SW480细胞株(48]。p . distasonis原油溶解产物和膜分数也被证明能降低疾病严重程度在全身的葡聚糖硫酸酯钠(DSS)结肠炎小鼠模型(60]。
我们并没有发现任何关联之间的隔离坚持肠道上皮细胞的能力,减弱LPS-induced炎症或加强上皮单层的完整性。然而,我们观察到细菌和肠上皮细胞之间的公司信息附件没有必要分离的菌株为发挥其抗炎活动作为他们的文化也浮在表面的减毒LPS-induced引发释放HT-29细胞系。期间我们没有解决细菌细胞的生存能力与HT-29细胞培养时间,但随着细菌上层清液也发现发挥抗炎作用,似乎效应分子的存在,而不是细菌生存本身是所需的活动。分泌的效应分子,如多糖,以及他们的交通工具,例如在omv,肠上皮细胞为宿主相互作用提供了一个解释在距离没有直接的物理相互作用[61年]。在肠道细菌omv也能穿过厚厚的一层粘液,甚至通过肠上皮细胞层。作为一个例子,omv分泌亚种发现了局部在宿主免疫细胞在肠道黏膜62年]。此外,b . vulgatus可以调节免疫细胞通过产生omv,通过粘蛋白层扩散和诱导树突状细胞TLR4和TLR2依赖的方式(49]。除了细菌效应分子,产生的短链脂肪酸(SCFAs)的细菌可以调节释放LPS-induced引发从上皮细胞63年]。
细菌效应分子的识别负责肠道内发挥抗炎作用仍然是一个悬而未决的问题。我们搜索的基因组拟杆菌和Parabacteroides隔离的wcfR基因,合成所需的保证(例如,PSA),但只有确定的基因组的同系物b . fragilis隔离。有趣的是,我们发现亚种spt基因(BT_0870)同系物的基因组显著相似b . caccae,b . intestinalis,b . fragilis和b . ovatus。的spt基因是参与鞘脂类的合成,调节bacteria-host交互的帮助下细菌omv [64年]。拟杆菌物种是已知的鞘脂类生产商在人类肠道但在多大程度上导致肠道内稳态仍然有待解决。殖民与sphingolipid-deficient无菌鼠亚种导致肠道炎症和神经酰胺池改变,表明细菌对肠道内稳态鞘脂类产生重大影响(24]。
拟杆菌和Parabacteroides种虫害包含有限合伙人的革兰氏阴性细菌,这被称为一个强有力的内毒素诱发强烈的促炎症反应的主机。然而,拟杆菌有限合伙人有截然不同的属性更多的有毒有限合伙人的肠杆菌科,包括大肠杆菌(65年]。的脂质中的一部分LPS结构负责endotoxicity影响配体亲和度TLR4-MD2复杂和下游NF-kB通路的激活(66年]。拟杆菌物种似乎拥有under-acylated,更少的有毒的脂质结构基于质谱分析显示表示“五”的存在和tetra-acylated脂质形式,而大肠杆菌拥有高促炎症,hexa-acylated脂质域(65年]。我们的WGS分析显示拟杆菌和Parabacteroides隔离本研究缺乏LpxL基因和LpxM需要构造一个hexa-acylated脂质a .相反的促炎症形式大肠杆菌有限合伙人,有限合伙人拟杆菌物种是免疫抑制65年,67年]。的确,拟杆菌有限合伙人可以调节的反应主要人类外周血单核细胞(PBMCs)大肠杆菌LPS刺激减少促炎细胞因子的生产,如TNF-a和il - 6 (65年]。此外,弱争胜拟杆菌有限合伙人被inflammation-reducing属性通过改善炎症免疫反应在一个实验性结肠炎小鼠模型68年]。因此,拟杆菌有限合伙人可能部分负责消炎观察在这个研究。
细菌性的spp,等Alistipes onderdonkii,b . fragilis和b . thetaiotamicron所示,monocolonised老鼠维持肠道内稳态通过促进结肠上皮内淋巴细胞(IEL) (51]。il - 6分泌通过IELs参与促进屏障功能和需要细菌MyD88信号。细菌性的物种增加il - 6在IELs体内而生产大肠杆菌控制,然而只有b . thetaiotamicron是增加显著。此外,b . fragilis和b . ovatus被证明缓解LPS-induced炎症在老鼠模型中通过减少TNF-α和增加细胞因子il - 10以及恢复Treg / Th17平衡[58]。有前景的结果关于细菌疗法治疗IBD的有效性是在最近的一项研究Ihekweazu et al。(2019)b . ovatus单一疗法改善结肠炎小鼠模型和增强上皮复苏更有效地比传统FMT [19]。小说的潜力隔离体内发挥抗炎作用还有待解决,但是以前的研究结果似乎在这方面鼓励。的共生体拟杆菌和Parabacteroides本研究分离与抗炎和上皮细胞增强能力是潜在的候选人关于bacteriotherapeutic应用程序旨在恢复肠道内稳态。分离的上皮加强行动展示了增强Caco-2单层完整性与媒介。然而,隔离能力防止不利影响的外部因素对上皮的完整性或侮辱后恢复完整没有研究,可能在将来的研究中得到解决。
总之,我们开发了一种高通量筛选的方法来分离菌株施加体外抗炎能力。在这项研究中,我们从健康的粪便分离供体拟杆菌和Parabacteroides物种减少的能力大肠杆菌LPS-induced引发生产从肠上皮细胞,增强上皮单层的完整性。这两个属性与隔离的粘附能力,然而花从分离介质的衰减能力持续使用表明,抗炎能力独立于细菌和上皮细胞之间的信息联系。在未来的研究中,我们的目标是识别潜在的效应分子并研究omv的角色参与抗炎能力,获得的WGS数据提供了一个理想的起点。潜在的细菌疗法治疗不同的条件正变得越来越明显,因此强调的重要性确定抗炎共生体,等拟杆菌spp。,他们的效应分子。
补充材料
以下是网上https://www.mdpi.com/2072-6643/12/4/935/s1,表S1:细菌的分离及其编码用于这项研究。全基因组测序的数据使用MiSeq (WGS)。,图S1:Estimates of evolutionary divergence by WGST (number of SNP differences per sequence) between the isolates belonging to the same species and previously published genomes (NCBI database). Bc =b . caccae(一),男朋友=b . fragilis(B), Bi =b . intestinalis(C),但是=b .均匀化(D), Bv =b . vulgatus(E),数字1 - 9指特定的隔离。数据p . distasonis和b . ovatus介绍了文本。补充excel文件。
作者的贡献
概念化R.S.V.K. K.H.;方法,R.S.V.K.,K.H.,D.L.,D.J.-S. and J.R.B.; validation, K.H., V.K., M.S., T.H., J.R.B., M.V. and R.B.; formal analysis, K.H., D.J.-S., D.L. and J.R.B.; writing—original draft preparation, K.H., R.S.; writing—review and editing, all authors; visualization, K.H., D.J.-S.; supervision, R.S., V.K. and D.M.E.; funding acquisition, R.S., V.K. All authors have read and agreed to the published version of the manuscript.
资金
这项研究是由芬兰科学院(格兰特nr。304490年和323156年为VK RS - 285632), Sigrid Juselius基金会FI (RS)高级研究员´s grant和微生物学和生物技术的博士项目,资助KH赫尔辛基大学。
确认
作者要感谢的粪便捐赠志愿者参与。
的利益冲突
作者宣称没有利益冲突。
引用
- Huttenhower c;Gevers d;骑士,r;Abubucker、美国结构、功能和健康的人类微生物组的多样性。自然2012年,486年,207 - 214。(谷歌学术搜索][CrossRef][绿色版本]
- 秦,j .;李,r;Raes, j .;Arumugam m;b . K.S.;Manichanh c;尼尔森,t;脑桥:;Levenez f;山田,t; et al. A human gut microbial gene catalogue established by metagenomic sequencing.自然2010年,464年59 - 65。(谷歌学术搜索][CrossRef][PubMed][绿色版本]
- 恩伯,P.J.;Hamady m;Yatsunenko t;Cantarel,文学士;邓肯,a;雷,镭射气;Sogin,马丁;琼斯,j•;罗伊,文学士学位;摩根大通(J.P. Affourtit; et al. A core gut microbiome in obese and lean twins.自然2009年,457年,480 - 484。(谷歌学术搜索][CrossRef][PubMed][绿色版本]
- 信仰,J.J.;Guruge J.L.;夏博诺,m;萨勃拉曼尼亚,美国;西多夫,h;古德曼,A.L.;克莱门特,J.C.;骑士,r;健康,交流;R.L. Leibel博士; et al. The long-term stability of the human gut microbiota.科学2013年,341年,1237439。(谷歌学术搜索][CrossRef][绿色版本]
- Salonen, a;Salojarvi, j .;拉赫蒂l;de Vos高家俊成人肠道微生物群是由核心分析深度和健康状况。中国。Microbiol。感染。2012年,18(5。4),16 - 20。(谷歌学术搜索][CrossRef][绿色版本]
- 冯·Hertzen l .;布鲁斯、b;Bienenstock称,j .;布拉斯m;Cani警察局;埃里克森,j .;Farkkila m;Haahtela t;Hanski i;Jenmalm,贝拉; et al. Helsinki alert of biodiversity and health.安。地中海。2015年,47,218 - 225。(谷歌学术搜索][CrossRef][绿色版本]
- Satokari, r .争议host-microbiota在炎症性肠病的关系,可以再次的敌人成为朋友?Scand。j .杂志。2015年,50,34-42。(谷歌学术搜索][CrossRef]
- Hiippala k;Jouhten h;龙凯宁,a;Hartikainen, a;Kainulainen诉;Jalanka, j .;Satokari, r .肠道共生体的潜力在加强肠屏障功能和减轻炎症。营养物质2018年,10。(谷歌学术搜索][CrossRef][绿色版本]
- 施罗德,影片;Backhed, f信号从肠道微生物群到遥远的器官在生理和疾病。Nat,地中海。2016年,22,1079 - 1089。(谷歌学术搜索][CrossRef]
- 弗兰克,向下;圣阿曼达A.L.;费尔德曼,R.A.;Boedeker,提到过;Harpaz:;速度,天然橡胶Molecular-phylogenetic表征人类炎性肠道疾病的微生物群落失衡。Proc。国家的。学会科学。美国2007年,104年,13780 - 13785。(谷歌学术搜索][CrossRef][绿色版本]
- Heidarian f;Alebouyeh m;Shahrokh,美国;Balaii h;Zali,核磁共振改变粪便细菌成分与疾病活动在炎症性肠病和IL8感应的范围。咕咕叫。Transl >,地中海。2019年,67年,每周。(谷歌学术搜索][CrossRef][PubMed]
- 周,y;智,f .低水平的拟杆菌在肠道微生物群与炎症性肠病:一个荟萃分析。生物医学。Int >,2016年,2016年,5828959。(谷歌学术搜索][CrossRef][PubMed][绿色版本]
- 奥图尔、毛重;Marchesi jr;山,c .下一代益生菌:频谱从益生菌biotherapeutics生活。Microbiol Nat。2017年,2,17057年。(谷歌学术搜索][CrossRef][PubMed]
- Cammarota g;Ianiro g;Tilg h;Rajilic-Stojanovic m;坎普,p;Satokari r;索科尔,h;Arkkila p;Pintus c;哈特,a; et al. European consensus conference on faecal microbiota transplantation in clinical practice.肠道2017年,66年,569 - 580。(谷歌学术搜索][CrossRef]
- 朱,w;冬天,M.G.;Byndloss M.X.;Spiga l;Duerkop,文学士学位;休斯,雌激素受体;Buttner l;德利马Romao大肠;贝伦特中一段;洛佩兹,c.a; et al. Precision editing of the gut microbiota ameliorates colitis.自然2018年,553年,208 - 211。(谷歌学术搜索][CrossRef]
- Moayyedi p;瑟特,M.G.;金,看法;Libertucci, j .;沃尔夫,m;Onischi c;阿姆斯特朗,d;马歇尔,J.K.;Kassam z;钟情,w; et al. Fecal Microbiota Transplantation Induces Remission in Patients with Active Ulcerative Colitis in a Randomized Controlled Trial.胃肠病学2015年,149年,102 - 109。(谷歌学术搜索][CrossRef][绿色版本]
- 雷森,净收益;f、s;van der machteld Spek M.J.;Tijssen J.G.;哈特曼,J.H.;Duflou, a;Lowenberg m;van den边缘,广义相对论;Mathus-Vliegen E.M.;德沃斯瓦特计; et al. Findings From a Randomized Controlled Trial of Fecal Transplantation for Patients With Ulcerative Colitis.胃肠病学2015年,149年,110 - 118。(谷歌学术搜索][CrossRef]
- 威尔逊,公元前;Vatanen t;Cutfield,至此;奥沙利文,J.M. Super-Donor现象在粪便微生物群移植。前面。细胞。感染。Microbiol。2019年,92。(谷歌学术搜索][CrossRef][绿色版本]
- Ihekweazu是;Fofanova T.Y.;Queliza k;Nagy-Szakal d;斯图尔特,C.J.;Engevik,硕士;Hulten kg。;Tatevian:;格雷厄姆,共晶;Versalovic, j .; et al. Bacteroides ovatus ATCC 8483 monotherapy is superior to traditional fecal transplant and multi-strain bacteriotherapy in a murine colitis model.肠道微生物2019年,1卷。(谷歌学术搜索][CrossRef][绿色版本]
- Staley c;凯撒,t;沃恩,安塞;Graiziger c;汉密尔顿,M.J.;Kabage抗干扰;Khoruts, a;Sadowsky, M.J.持久长期细菌移植后封装粪便微生物群移植治疗艰难梭状芽胞杆菌感染。MBio2019年,10。(谷歌学术搜索][CrossRef][绿色版本]
- Jalanka, j .;马提拉,大肠;Jouhten h;哈特曼,j .;德沃斯瓦特计;Arkkila p;Satokari, r .腔的长期影响和粘膜微生物群,通常获得类群在粪便微生物群移植梭状芽胞杆菌对复发性感染。BMC地中海。2016年,14,155年。(谷歌学术搜索][CrossRef][PubMed][绿色版本]
- Zitomersky N.L.;科因,M.J.;康斯托克,L.E.患病率的纵向分析,维护和IgA应对订单的物种在人类肠道细菌性的。感染。Immun。2011年,79年,2012 - 2020。(谷歌学术搜索][CrossRef][PubMed][绿色版本]
- Wexler A.G.;古德曼A.L.内幕的角度:拟杆菌作为窗口的微生物。Microbiol Nat。2017年,2,17026年。(谷歌学术搜索][CrossRef][PubMed][绿色版本]
- 布朗,E.M.;柯,x;希区柯克,d;Jeanfavre,美国;Avila-Pacheco, j .;醒来时,t;亚瑟,杰;Fornelos:;海姆,c;电子艺界Franzosa; et al. Bacteroides-Derived Sphingolipids Are Critical for Maintaining Intestinal Homeostasis and Symbiosis.细胞宿主细菌2019年,25,668 - 680。(谷歌学术搜索][CrossRef]
- Mazmanian”栏目;圆的,评论;卡斯帕,D.L.微生物共生因素阻止肠道炎性疾病。自然2008年,453年,620 - 625。(谷歌学术搜索][CrossRef][PubMed][绿色版本]
- 沈,y;Giardino Torchia,马丁;劳森,G.W.;卡普,中一段;阿什维尔,J.D.;Mazmanian,”栏目外膜囊泡的人类共生的调解免疫调控和疾病的保护。细胞宿主细菌2012年,12,509 - 520。(谷歌学术搜索][CrossRef][绿色版本]
- Waidmann m;Bechtold o .;弗里克,j.s;莱尔,H.A.;舒伯特,美国;Dobrindt,美国;吕弗勒,j .;Bohn大肠;Autenrieth, I.B.拟杆菌vulgatus预防大肠coli-induced结肠炎在无菌的interleukin-2-deficient老鼠。胃肠病学2003年,125年,162 - 177。(谷歌学术搜索][CrossRef]
- Satokari r;马提拉,大肠;Kainulainen诉;Arkkila,体育简单粪便准备和冷冻剂的功效在粪便微生物群移植复发感染梭状芽胞杆菌——一项观察性队列研究。滋养品。杂志。其他。2015年,41,46-53。(谷歌学术搜索][CrossRef]
- Kainulainen诉;唐,y;斯皮尔曼、t;Kilpinen,美国;Reunanen, j .;莎丽服、体育;Satokari, r .犬分离嗜酸乳杆菌LAB20坚持肠道上皮细胞,变弱LPS-induced引发肠上皮细胞体外分泌。BMC Microbiol。2015年,154。(谷歌学术搜索][CrossRef][绿色版本]
- 发送方,r;福克斯,美国;米洛,r .修正估计人类和细菌细胞的数量。公共科学图书馆杂志。2016年,14,e1002533。(谷歌学术搜索][CrossRef][绿色版本]
- 罗尔夫,进食;昂特热,D.J.;坎贝尔,B.J.;巴雷特,j.t因素相关的氧厌氧细菌公差。达成。环绕。Microbiol。1978年,36,306 - 313。(谷歌学术搜索][CrossRef][PubMed][绿色版本]
- Papanicolas L.E.;Choo J.M.;王,y;梁,L.E.X.;科斯特洛,……;戈登,D.L.;Wesselingh S.L.;罗杰斯,粪便移植G.B.细菌生存能力:细菌生存?EBioMedicine2019年,41,509 - 516。(谷歌学术搜索][CrossRef][PubMed][绿色版本]
- 基础,聚合度;Tuckova l;Farre,硕士;Iwase t;拥抱我;Tlaskalova-Hogenova、h . CD14表达和释放可溶性CD14的人类肠道上皮细胞体外:上皮细胞的脂多糖活化再现。感染。Immun。2001年,69年,3772 - 3781。(谷歌学术搜索][CrossRef][绿色版本]
- 许,R.Y.;陈,h;Spicer J.D.;卢梭,贝拉;Giannias b;卢梭,美国;费里,L.E. LPS-induced TLR4信号在人类大肠癌细胞增加beta1 integrin-mediated细胞粘附和肝脏转移。癌症Res。2011年,71年,1989 - 1998。(谷歌学术搜索][CrossRef][PubMed][绿色版本]
- Van De机器人瓦力,j .;亨德里克斯,a;罗密,b;Larondelle y;施耐德,Y.J. Caco-2细胞炎症参数:刺激效果自然、浓度、组合和细胞分化。毒理学2010年,24,1441 - 1449。(谷歌学术搜索][CrossRef]
- Kainulainen诉;Reunanen, j .;Hiippala k;Guglielmetti,美国;Vesterlund,美国;Palva, a;Satokari, r . BopA没有重要作用在双歧杆菌的粘附bifidum肠道上皮细胞,细胞外基质蛋白,和粘液。达成。环绕。Microbiol。2013年,79年,6989 - 6997。(谷歌学术搜索][CrossRef][绿色版本]
- Reunanen, j .;Kainulainen诉;Huuskonen l;Ottman:;茨,c;Huhtinen h;德沃斯瓦特计;Satokari, r . Akkermansia muciniphila坚持肠上皮细胞,增强上皮细胞层的完整性。达成。环绕。Microbiol。2015年,81年,3655 - 3662。(谷歌学术搜索][CrossRef][绿色版本]
- 伊达尔戈,I.J.;Raub T.J.;波哈特,保留时间表征人类结肠癌细胞系(Caco-2)作为一个模型系统肠道上皮通透性。胃肠病学1989年,96年,736 - 749。(谷歌学术搜索][CrossRef]
- 谢长廷,C.Y.;大阪、t;本森山,大肠;目前为止,y;菊池,j .;Tsuneda,美国加强肠上皮细胞紧密连接的双歧杆菌bifidum。杂志。代表。2015年,3。(谷歌学术搜索][CrossRef][绿色版本]
- Klingberg杰;Pedersen M.H.;Cencic, a;布德博应用transepithelial电阻的测量肠道上皮细胞层评价益生菌的活动。达成。环绕。Microbiol。2005年,71年,7528 - 7530。(谷歌学术搜索][CrossRef][绿色版本]
- Sahl J.W.;登月舱,d;特拉维斯,j .;思楚普J.M.;Gillece J.D.;阿齐兹,m;Driebe E.M.;德雷斯,陷入;菲尔特希克斯;威廉姆森,C.H.D.; et al. NASP: An accurate, rapid method for the identification of SNPs in WGS datasets that supports flexible input and output formats.活细胞。染色体组。2016年,2,e000074。(谷歌学术搜索][CrossRef][PubMed]
- 李,h;杜宾,r .快速和准确的短阅读符合burrows - wheeler变换。生物信息学2009年,25,1754 - 1760。(谷歌学术搜索][CrossRef][PubMed][绿色版本]
- 麦肯纳,a;汉娜,m;银行,大肠;Sivachenko, a;Cibulskis k;Kernytsky, a;盖瑞,k;Altshuler d;盖伯瑞尔,美国;戴利,m; et al. The Genome Analysis Toolkit: A MapReduce framework for analyzing next-generation DNA sequencing data.基因组Res。2010年,20.,1297 - 1303。(谷歌学术搜索][CrossRef][PubMed][绿色版本]
- Delcher A.L.;Phillippy, a;卡尔顿,j .;扎尔茨贝格,S.L.快速算法对大规模基因组比对和比较。核酸Res。2002年,30.,2478 - 2483。(谷歌学术搜索][CrossRef][绿色版本]
- 阮,L.T.;施密特,H.A.;冯·Haeseler a;明,B.Q. IQ-TREE:一个快速和有效的随机算法估计最大似然的发展史。摩尔。杂志。另一个星球。2015年,32,268 - 274。(谷歌学术搜索][CrossRef]
- 库马尔,美国;Stecher g;(k . MEGA7:分子进化遗传学分析7.0版本的更大的数据集。摩尔。杂志。另一个星球。2016年,33,1870 - 1874。(谷歌学术搜索][CrossRef][绿色版本]
- Stamatakis RAxML version 8:系统发育分析的工具和大型的发展史后分析。生物信息学2014年,30.,1312 - 1313。(谷歌学术搜索][CrossRef]
- Koh G.Y.;凯恩,a;李,k;徐,问:;吴,x;罗珀,j .;梅森,J.B.;Crott, J.W. Parabacteroides distasonis变弱toll样受体4信号和Akt激活和块结肠肿瘤形成的高脂肪饮食azoxymethane-treated老鼠。Int。j .癌症2018年。(谷歌学术搜索][CrossRef][绿色版本]
- Maerz J.K.;Steimle, a;兰格,a;本德,a;一定要,b;弗里克,j.s外膜囊泡起泡有助于b vulgatus mpk-mediated免疫反应沉默。肠道微生物2018年,9,1 - 12。(谷歌学术搜索][CrossRef][绿色版本]
- 王,k;廖,m;周:;包,l;马,k;郑,z;王,y;刘,c;王,w;王,j .; et al. Parabacteroides distasonis Alleviates Obesity and Metabolic Dysfunctions via Production of Succinate and Secondary Bile Acids.细胞的代表。2019年,26,222 - 235。(谷歌学术搜索][CrossRef][绿色版本]
- 库恩·;舒尔茨,莫莱森;Regner,再见;一口油井为el瑞,塞维,Hendrickson, J.D.;梅塔,g;惠特尼,A.K.;红外光谱、d;Ohri:;罗伯逊,着力点; et al. Bacteroidales recruit IL-6-producing intraepithelial lymphocytes in the colon to promote barrier integrity.粘膜Immunol。2018年,11,357 - 368。(谷歌学术搜索][CrossRef][PubMed][绿色版本]
- 西北点;恩斯特,上面;蔡,J.H.;威尔逊,C.B.;米勒,S.I.人类toll样受体4识别寄主专一性的有限合伙人的修改。Immunol Nat。2002年,3,354 - 359。(谷歌学术搜索][CrossRef][PubMed]
- Polissi, a;Sperandeo, p .大肠杆菌脂多糖出口途径:结构,组织和调节涡轮机械的装配。3月的药物2014年,12,1023 - 1042。(谷歌学术搜索][CrossRef][PubMed][绿色版本]
- De Freitas M.B.;Moreira E.A.M.;Tomio c;莫雷诺,Y.M.F.;Daltoe F.P.;巴博萨,大肠;路德维希否决权:;Buccigrossi诉;Guarino, a改变肠道微生物群组成,抗生素治疗和肠道炎症在囊性纤维化的儿童和青少年。《公共科学图书馆•综合》2018年,13,e0198457。(谷歌学术搜索][CrossRef]
- Gevers d;Kugathasan,美国;Denson,洛杉矶;Vazquez-Baeza y;范Treuren w;任,b;Schwager大肠;骑士,d;歌,中华民国;Yassour m; et al. The treatment-naive microbiome in new-onset Crohn’s disease.细胞宿主细菌2014年,15,382 - 392。(谷歌学术搜索][CrossRef][PubMed][绿色版本]
- Paramsothy,美国;尼尔森,美国;卡姆,硕士;Deshpande N.P.;信仰,J.J.;克莱门特,J.C.;Paramsothy r;沃尔什,抗干扰;van den Bogaerde, j .;塞缪尔·d·; et al. Specific Bacteria and Metabolites Associated With Response to Fecal Microbiota Transplantation in Patients With Ulcerative Colitis.胃肠病学2019年,156年,1440 - 1454。(谷歌学术搜索][CrossRef][绿色版本]
- Delday m;穆德,我;洛根,E.T.;格兰特,g .叫多形拟改善结肠炎症在克罗恩病的临床前模型。Inflamm。肠道说。2019年,25,85 - 96。(谷歌学术搜索][CrossRef][绿色版本]
- 棕褐色,h;赵,j .;张,h;翟,问:;陈威小说脆弱拟杆菌、拟杆菌菌株ovatus缓解LPS-induced炎症小鼠。达成。Microbiol。Biotechnol。2019年,103年,2353 - 2365。(谷歌学术搜索][CrossRef]
- Truong D.T.;邰蒂,a;Pasolli大肠;Huttenhower c;Segata:微生物strain-level人口从基因组结构和基因多样性。基因组Res。2017年,27,626 - 638。(谷歌学术搜索][CrossRef]
- Kverka m;Zakostelska z;Klimesova k;索科尔,d;Hudcovic t;Hrncir t;罗斯曼,p;穆雷查克,j .;Kopecny, j .;一直对E.F.; et al. Oral administration of Parabacteroides distasonis antigens attenuates experimental murine colitis through modulation of immunity and microbiota composition.中国。Exp Immunol。2011年,163年,250 - 259。(谷歌学术搜索][CrossRef]
- Schwechheimer c;期,M.J.从革兰氏阴性细菌外膜囊泡:生源论和功能。启Microbiol Nat。2015年,13,605 - 619。(谷歌学术搜索][CrossRef][PubMed][绿色版本]
- 希基,c.a;库恩·;Donermeyer D.L.;波特,自备;金,c;卡梅伦,电子;荣格,h;Kaiko,通用电气公司;Wegorzewska m;Malvin N.P.; et al. Colitogenic Bacteroides thetaiotaomicron Antigens Access Host Immune Cells in a Sulfatase-Dependent Manner via Outer Membrane Vesicles.细胞宿主细菌2015年,17,672 - 680。(谷歌学术搜索][CrossRef][PubMed][绿色版本]
- Asarat m;Vasiljevic t;Apostolopoulos诉;Donkor, o .短链脂肪酸调节分泌引发的人类肠道上皮细胞株体外。Immunol。Investig。2015年,44,678 - 693。(谷歌学术搜索][CrossRef][PubMed]
- 挑夫,S.L.;一口油井为el瑞,约翰逊,雷,右眼鞘脂类在host-microbial交互。咕咕叫。当今。Microbiol。2018年,43,92 - 99。(谷歌学术搜索][CrossRef]
- D 'Hennezel大肠;Abubucker,美国;墨菲,润滑;卡伦卢铁荣总脂多糖的人类肠道微生物组沉默toll样受体信号。mSystems2017年,2。(谷歌学术搜索][CrossRef][绿色版本]
- Nijland r;Hofland t;范Strijp,正当识别有限合伙人的TLR4:潜在的抗炎治疗。3月的药物2014年,12,4260 - 4273。(谷歌学术搜索][CrossRef][绿色版本]
- Vatanen t;公元Kostic;D 'hennezel大肠;Siljander h;电子艺界Franzosa;Yassour m;Kolde r;Vlamakis h;亚瑟,杰;Hamalainen点; et al. Variation in Microbiome LPS Immunogenicity Contributes to Autoimmunity in Humans.细胞2016年,165年,842 - 853。(谷歌学术搜索][CrossRef][绿色版本]
- Steimle, a;米歇利斯,l;Di Lorenzo f;Kliem t;Munzner t;Maerz J.K.;谢弗,a;兰格,a;Parusel r;Gronbach k; et al. Weak Agonistic LPS Restores Intestinal Immune Homeostasis.摩尔。其他。2019年。(谷歌学术搜索][CrossRef][绿色版本]
图1所示。的高通量方法筛选抗炎细菌从不同的步骤和结果。

图2。估计进化趋异的全基因组单核苷酸多态性(SNP)输入(WGST)(每个序列的SNP数量差异)之间的隔离属于同一个物种和以前公布的基因组(NCBI数据库)。面板(一个)是p . distasonis(Pd)和面板(B)是b . ovatus(Bo)。
图2。估计进化趋异的全基因组单核苷酸多态性(SNP)输入(WGST)(每个序列的SNP数量差异)之间的隔离属于同一个物种和以前公布的基因组(NCBI数据库)。面板(一个)是p . distasonis(Pd)和面板(B)是b . ovatus(Bo)。

图3。坚持的隔离Caco-2 HT-29细胞系和肠道粘液。Pd =p . distasonis公元前=b . caccae男朋友=b . fragilisBi =b . intestinalisBu =b .均匀化Bv =b . vulgatus博=b . ovatus。数字1 - 9指的是特定的隔离。符号+显示隔离的能力减弱LPS-induced引发释放HT-29细胞系。
图3。坚持的隔离Caco-2 HT-29细胞系和肠道粘液。Pd =p . distasonis公元前=b . caccae男朋友=b . fragilisBi =b . intestinalisBu =b .均匀化Bv =b . vulgatus博=b . ovatus。数字1 - 9指的是特定的隔离。符号+显示隔离的能力减弱LPS-induced引发释放HT-29细胞系。

图4。使用了隔离介质的衰减能力。、文化上层清液。岐阜厌氧培养基(GAM)和本人5媒体以及只有有限合伙人作为控制。有限合伙人控制代表100%引发释放HT-29细胞。½和¼稀释试验中使用的媒体。Pd =p . distasonis公元前=b . caccae男朋友=b . fragilisBi =b . intestinalisBu =b .均匀化Bv =b . vulgatus博=b . ovatus。数字1 - 9是指特定的隔离。
图4。使用了隔离介质的衰减能力。、文化上层清液。岐阜厌氧培养基(GAM)和本人5媒体以及只有有限合伙人作为控制。有限合伙人控制代表100%引发释放HT-29细胞。½和¼稀释试验中使用的媒体。Pd =p . distasonis公元前=b . caccae男朋友=b . fragilisBi =b . intestinalisBu =b .均匀化Bv =b . vulgatus博=b . ovatus。数字1 - 9是指特定的隔离。

图5。在transepithelial隔离电阻的影响(TER) Caco-2单层。大肠杆菌作为一个消极的控制。结果显示平均额,价值变化(Ω/厘米2在24小时内)和标准偏差的三个技术复制(平行威尔斯)。星号表示明显不同(p< 0.05)值比未经处理的Caco-2单层膜(媒介控制)。*p< 0.05,* *p< 0.01,* * *p< 0.001。Pd =p . distasonis公元前=b . caccaeBi =b . intestinalisBu =b .均匀化Bv =b . vulgatus博=b . ovatus。数字1 - 9指的是特定的隔离。
图5。在transepithelial隔离电阻的影响(TER) Caco-2单层。大肠杆菌作为一个消极的控制。结果显示平均额,价值变化(Ω/厘米2在24小时内)和标准偏差的三个技术复制(平行威尔斯)。星号表示明显不同(p< 0.05)值比未经处理的Caco-2单层膜(媒介控制)。*p< 0.05,* *p< 0.01,* * *p< 0.001。Pd =p . distasonis公元前=b . caccaeBi =b . intestinalisBu =b .均匀化Bv =b . vulgatus博=b . ovatus。数字1 - 9指的是特定的隔离。


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
表1。的能力拟杆菌和Parabacteroides全身的隔离减弱脂多糖(LPS)引发释放HT-29细胞系。
| 隔离 | 显著的减少引发* / 总数衰减实验 |
%下降引发释放 相比,有限合伙人控制 |
衰减 能力# |
|---|---|---|---|
| Pd1 | 1/4 | 7-53 | - - - - - - |
| Pd2 | 4/5 | 35 - 61 | + |
| Pd3 | 3/3 | 27-39 | + |
| Pd4 | 4/6 | 23-59 | + |
| Pd5 | 0/3 | 划分的 | - - - - - - |
| Pd6 | 0/3 | 15 - 23 | - - - - - - |
| 群体Bc1 | 4/6 | 12-44 | + |
| Bc2 | 3/4 | 21-46 | + |
| Bc3 | 3/5 | 33-48 | + |
| 男朋友 | 0/3 | 猴 | - - - - - - |
| Bi1 | 3/3 | 21 - 72 | + |
| Bi2 | 3/4 | 25 - 66 | + |
| Bi3 | 3/4 | 27-51 | + |
| 部 | 4/6 | 34-59 | + |
| Bv1 | 3/4 | 26 39 | + |
| Bv2 | 3/3 | 23-36 | + |
| Bv3 | 5/9 | 39-59 | + |
| Bv4 | 3/3 | 批准 | + |
| Bv5 | 3/3 | 41-55 | + |
| Bo1 | 2/5 | 5-58 | - - - - - - |
| Bo2 | 4/8 | 0-39 | - - - - - - |
| Bo3 | 4/6 | 30 - 50 | + |
| Bo4 | 3/4 | 一 | + |
| Bo5 | 3/7 | 0-39 | - - - - - - |
| Bo6 | 0/3 | 17-26 | - - - - - - |
| Bo7 | 3/7 | 0-28 | - - - - - - |
| Bo8 | 2/5 | 0-54 | - - - - - - |
| Bo9 | 1/5 | 0-45 | - - - - - - |
*重要(p< 0.05)减少引发释放和控制,即。,有限合伙人stimulation without a prior exposure to the studied isolate. #no attenuation or seldomly detected (−), attenuation constantly detected (+). Pd =p . distasonis公元前=b . caccae男朋友=b . fragilisBi =b . intestinalisBu =b .均匀化Bv =b . vulgatus博=b . ovatus。数字1 - 9指的是特定的隔离。#隔离显著(p≤0.05)减少引发释放在至少三个独立的实验,在多数的实验中被认为有衰减能力(+)。
作者©2020年。被许可方MDPI,瑞士巴塞尔。这篇文章是一个开放存取物品分发的条款和条件下知识共享归属(CC)许可证(http://creativecommons.org/licenses/by/4.0/)。
分享和引用
MDPI和ACS风格
Hiippala k;Kainulainen诉;Suutarinen m;海尼、t;鲍尔斯,jr;Jasso-Selles d;登月舱,d;情人节,m;巴恩斯,r;Engelthaler D.M.; Satokari, R. Isolation of Anti-Inflammatory and Epithelium Reinforcing拟杆菌和Parabacteroides种虫害从健康的粪便捐赠。营养物质2020年,12,935年。https://doi.org/10.3390/nu12040935
AMA风格
Hiippala K, Kainulainen V, Suutarinen M,海尼T,小凉亭,Jasso-Selles D,登月舱D,情人节M,巴恩斯R, Engelthaler DM, Satokari R .隔离的抗炎和上皮加强拟杆菌和Parabacteroides种虫害从健康的粪便捐赠。营养物质。2020;12 (4):935。https://doi.org/10.3390/nu12040935
芝加哥/ Turabian风格Hiippala,佳兆业集团、维拉Kainulainen Maiju Suutarinen,图海尼,茱莲妮r·鲍尔斯Daniel Jasso-Selles达登月舱,迈克尔•瓦伦丁Riley巴恩斯和David m . Engelthaler Reetta Satokari》2020。“孤立的抗炎和上皮加固拟杆菌和Parabacteroides种虫害从健康的粪便捐赠”营养物质12,没有。4:935。https://doi.org/10.3390/nu12040935
注意,从2016年的第一期,这个期刊使用的文章而不是页面号码。看到更多的细节在这里。