条文本

摘要
客观的非酒精性脂肪肝(NAFLD)相关的肝细胞癌(HCC)是世界范围内日益加重的医疗负担。我们研究了膳食胆固醇通过调节肠道菌群及其代谢物在驱动NAFLD-HCC中的作用。
设计以高脂/高胆固醇(HFHC)、高脂/低胆固醇或正常饲料喂养C57BL/6窝雄猫14个月。给hfhc喂养的小鼠服用降胆固醇药物阿托伐他汀。无菌小鼠移植不同饲料小鼠的粪便,以确定胆固醇调节菌群在NAFLD-HCC中的直接作用。肠道菌群采用16S rRNA测序,血清代谢产物采用液相色谱-质谱(LC-MS)代谢组学分析。对59例高胆固醇血症患者和39例健康对照者的粪便微生物成分进行了检测。
结果高胆固醇导致小鼠脂肪变性、脂肪性肝炎、纤维化和最终肝细胞癌的顺序进展,并伴有胰岛素抵抗。胆固醇诱导的NAFLD-HCC的形成与肠道菌群失调有关。在脂肪变性、脂肪性肝炎和肝细胞癌的各个阶段,微生物群的组成明显集中。脱磷孤菌属,Mucispirillum Anaerotruncus而且Desulfovibrionaceae增加的顺序;而双歧杆菌属而且拟杆菌在hfhc喂养的小鼠中减少,这在人类高胆固醇血症患者中得到证实。膳食胆固醇引起肠道细菌代谢产物的改变,包括牛磺胆酸增加和3-吲哚丙酸减少。无菌小鼠灌胃经HFHC喂养的小鼠粪便后表现为肝脏脂质堆积、炎症和细胞增殖。此外,阿托伐他汀恢复了胆固醇诱导的肠道菌群紊乱,并完全阻止了NAFLD-HCC的发展。
结论膳食胆固醇通过诱导小鼠肠道菌群和代谢物的改变,驱动NAFLD-HCC的形成。胆固醇抑制疗法和肠道菌群控制可能是预防NAFLD-HCC的有效策略。
- 饮食因素
- 脂肪肝
- 非酒精性脂肪肝炎
- 肠道微生物
这是一篇开放获取的文章,按照创作共用署名非商业性(CC BY-NC 4.0)许可发布,该许可允许其他人以非商业性的方式发布、混编、改编、构建本作品,并以不同的条款授权他们的衍生作品,前提是原创作品被正确引用,给予适当的荣誉,任何更改都被注明,且使用是非商业性的。看到的:http://creativecommons.org/licenses/by-nc/4.0/.
来自Altmetric.com的统计
本研究的意义
关于这个问题,我们已经知道了什么?
非酒精性脂肪肝(NAFLD)相关的肝细胞癌(HCC)是世界范围内日益加重的医疗负担。
胆固醇是一种主要的脂毒分子。
新的发现是什么?
高脂高胆固醇饮食(HFHC)自发和依次诱导雄性小鼠脂肪肝、脂肪性肝炎、纤维化和NAFLD-HCC的发生,而高脂低胆固醇饮食仅诱导雄性小鼠肝脏脂肪变性。
膳食胆固醇诱导的NAFLD-HCC的形成与肠道菌群失调有关。在NAFLD-HCC形成的各个阶段,菌群组成发生了变化:脱磷孤菌属,Mucispirillum Anaerotruncus而且Desulfovibrionaceae依次增加;而双歧杆菌属而且拟杆菌在hfhc喂养的小鼠中减少,这在人类高胆固醇血症患者中得到证实。
在NAFLD-HCC中发现肠道细菌代谢物的改变,包括血清牛磺胆酸(TCA)增加和3-吲哚丙酸(IPA)减少。IPA抑制胆固醇诱导的脂质积累和细胞增殖,而TCA加剧胆固醇诱导的甘油三酯在人正常永生化肝细胞的积累。
用hfhc喂养的小鼠粪便灌胃的无菌小鼠表现出肝脏脂质积累、炎症和细胞增殖,与供体小鼠表型一致。
抗胆固醇治疗恢复了膳食胆固醇诱导的肠道菌群紊乱,并完全阻止了NAFLD-HCC的形成。
在可预见的未来,它会对临床实践产生怎样的影响?
我们的研究结果表明,抗胆固醇药物和操纵肠道菌群可能是预防NAFLD-HCC的有效策略。
简介
非酒精性脂肪性肝病(NAFLD)是代谢综合征的肝脏表现,包括从单纯脂肪变性到非酒精性脂肪性肝炎(NASH)的一系列肝脏病理。1NASH可进展为肝硬化、终末期肝衰竭和肝细胞癌(HCC)。2目前,NAFLD是全球发病率的主要原因和医疗负担。以人群为基础的大型队列研究表明,与肝炎的2.5倍相比,NAFLD-HCC的患病率在过去十年中增加了4倍,使其成为肝移植中增长最快的指征。3.脂肪毒性驱动NASH、纤维化/肝硬化,甚至肝癌的进展。4在肝脂类中,胆固醇被认为是NASH发展的主要脂毒分子。4肝脏是调节全身胆固醇内稳态的中心。在NASH的人类和实验模型中都证实了肝脏胆固醇稳态的异常。5个6我们已经发现,角鲨烯环氧化酶,一种胆固醇生物合成的速率限制酶,驱动NAFLD-HCC的发展。7膳食胆固醇对血浆和肝脏胆固醇稳态有重要影响。5虽然已经报道了饮食胆固醇对NASH进展的影响,5 8长期胆固醇治疗在自发性和进行性NAFLD-HCC发展中的作用和致病基础尚不清楚。
肠道菌群与宿主具有共生关系,通过代谢包括大肠胆固醇在内的膳食成分,为宿主提供营养和能量。9一些研究表明,肠道微生物群是促进NAFLD发展及其进展为NAFLD - hcc的环境因素。10,11微生物衍生的代谢物,如胆汁酸、短链脂肪酸和三甲胺及其影响的信号通路可能有助于NAFLD的发展。12日13特别是3-(4-羟基苯基)乳酸盐,一种来自肠道微生物组的代谢物,对肝脏脂肪变性和纤维化具有共同的基因效应。14此外,饮食胆固醇对肠道菌群的影响也有报道。15然而,肠道菌群失调是否是饮食胆固醇诱导的NASH和NAFLD-HCC进展的原因或影响尚不清楚。
本研究旨在确定膳食胆固醇在NAFLD-HCC发展中的作用及其相关分子机制。我们发现,膳食胆固醇通过胆固醇诱导的肠道菌群变化和代谢组学改变,导致NAFLD-HCC的自发形成。胆固醇抑制恢复了肠道菌群,并完全阻止了NAFLD-HCC的发展。
材料和方法
动物和饮食
雄性C57BL/6野生型窝伴侣(8周龄)自由饲喂正常饲料(NC、18%脂肪、58%碳水化合物、24%蛋白质、0%胆固醇)、高脂肪/低胆固醇饲料(hfc、43.7%脂肪、36.6%碳水化合物、19.7%蛋白质、0.013%胆固醇)或高脂肪/高胆固醇饲料(HFHC、43.7%脂肪、36.6%碳水化合物、19.7%蛋白质、0.203%胆固醇)(Specialty Feeds, Glen Forrest, WA) 14个月。各组分别饲喂HFLC和HFHC 3、8、10和12个月(每组n=8 ~ 19只)。在其他实验中,在开始饮食的7个月后,给hfhc喂养的小鼠服用阿托伐他汀(20 mg/kg),并继续7个月(每组n=10)。小鼠被保持12/12小时的光/暗循环。在实验终点,小鼠禁食并收集血清/组织。16记录体重和内脏脂肪重量。肝脏被迅速切除并称重。评估表面结节的存在和尺寸。从组织学上看,肝细胞癌是由粗大结节和组织学上明显的结节证实的。肝肿瘤被分离出来,快速冷冻在液氮中,并储存在−80°C,以供进一步实验。取小鼠粪便标本进行细菌16S rRNA基因测序和灌胃。
C57BL/6雄性无菌小鼠(7周)在重庆第三军医大学实验动物学系繁殖。将成年小鼠(8周龄)分为3组(每组11 ~ 28只),分别于0月龄和7月龄分别灌胃2次,灌胃小鼠粪便分别取自常规小鼠喂养的NC、HFLC、HFHC或经阿托伐他汀治疗的HFHC小鼠,共14个月。简单地说,1克粪便样本均质于5毫升磷酸盐缓冲盐水(PBS)中。用200 μL的悬液灌胃移植受体小鼠。每组小鼠在移植后8个月、10个月或14个月随机抽取并处死。
中提供了其他方法在线补充文件1.
结果
饮食胆固醇驱动NAFLD-HCC的自发发展
为了研究膳食胆固醇在脂肪变性、NASH、纤维化和随后的NAFLD-HCC的发展过程中的作用,小鼠被喂以HFHC,17HFLC或数控。在不同时间点(3、8、10、12、14个月)监测血清甲胎蛋白(AFP,肝癌标志物)水平。高脂喂养小鼠的甲胎蛋白水平在第10个月时(107.1±127.9 ng/mL)升高,在第12个月时(151.6±129.3 ng/mL)和第14个月时(174.2±203.1 ng/mL)进一步升高,而高脂喂养小鼠的甲胎蛋白水平在第14个月时分别为50.9±7.5 ng/mL和59.7±20.9 ng/mL。图1一个).第14个月时,MRI扫描显示hfhc喂养的小鼠有肝脏肿瘤,但hflc喂养或nc喂养的小鼠没有(图1 b).然后在第14个月时收获小鼠。68%(13/19)的高脂高脂喂养的小鼠发现了肝肿瘤,但在高脂高脂或NC喂养的小鼠中没有发现(图1 c).肝脏切片的组织学检查证实所有肝脏肿瘤均为肝细胞癌(图1 c),平均每只小鼠有2.7±2.6个肿瘤细胞,最大肿瘤直径为4.1±5.0 mm。与hflc喂养的小鼠相比,hflc喂养的小鼠肝脏切片有明显更多的Ki-67阳性细胞,表明hflc喂养的小鼠细胞增殖增加(图1 c).这些结果表明,饮食中的胆固醇可以自发诱导NAFLD-HCC的形成。

胆固醇诱导C57BL/6小鼠自发性肝癌形成。(A)高脂饲料3、8、10、12、14个月和NC、HFLC饲料14个月小鼠血清甲胎蛋白水平;(B)肝脏MRI, (C)饲喂NC、HFLC和HFHC 14个月小鼠肝脏代表性大体形态、代表性显微镜特征和Ki67染色免疫组化图,Ki-67按以下标准评分%细胞染色),1 (11%-30%细胞染色),2 (30%-50%细胞染色)或3 (>50%细胞染色);(D)饲喂NC、HFLC和HFHC 14个月小鼠的体重、内脏脂肪、肝脏重量、肝重比,(E)血清胆固醇水平、肝脏游离胆固醇、肝脏胆固醇酯含量、糖耐量试验和空腹胰岛素水平。* * * p < 0.05, p < 0.01, * * * p < 0.001。据法新社,甲胎蛋白;数控,正常的食物;HFLC、高脂肪、低胆固醇的饮食;饲料、高脂肪、高胆固醇的饮食;H&E,苏木精和伊红。
在HCC形成的第14个月,与对照组相比,HFHC喂养的小鼠体重、内脏脂肪、肝脏重量和肝重比均显著增加(图1 d).喂食hflc的小鼠体重、内脏脂肪和肝脏重量均有所增加(图1 d).与hflc或nc喂养的小鼠相比,hfhc喂养的小鼠血清胆固醇、肝脏游离胆固醇和胆固醇酯、葡萄糖不耐受和空腹胰岛素显著增加(图1 e).与NC相比,HFLC喂养的小鼠也观察到葡萄糖不耐受和空腹胰岛素增强(图1 e).
高胆固醇饮食喂养14个月的小鼠出现NASH和纤维化
进一步的肝脏切片检查显示,在喂高脂hc 14个月的小鼠的肝细胞癌邻近肝组织和非肝细胞癌肝组织中存在以脂肪变性和小叶炎症为特征的脂肪性肝炎,而只在喂高脂hc的小鼠中观察到脂肪变性(图2一个,在线补充图S1).与组织学炎症一致,hfhc喂养的小鼠血清谷丙转氨酶(ALT) (p<0.01)和天冬氨酸转氨酶(AST) (p<0.01)水平显著高于HFLC或NC喂养的小鼠(p<0.01)。图2 b).血清和肝脏促炎细胞因子包括IL-6、IL-1α和IL-1β (图2 c, D)及促炎因子(在线补充图S2A-B细胞因子谱分析和酶联免疫吸附测定结果显示,hfhc喂养的小鼠中)的表达增加。RNA测序分析表明,与hflc喂养的小鼠相比,hflc喂养的14月龄小鼠的肝组织中重要的与nash相关的促炎细胞因子,包括Cx3cl1, Mcp1, Cxcl10, Mip1β, Mip1α, Ccl5, Cxcl16和Tnfα,显著上调。图2 d).hfhc喂养的小鼠出现严重的纤维化损伤,胶原蛋白分布区域明显增多(图2 e)、肝脏羟脯氨酸测定胶原蛋白含量(图2 f)和肝星状细胞激活,其表现为α-平滑肌肌动蛋白(α-SMA) mRNA和蛋白水平的增加(图2 e).肝脏氧化应激检查显示,氧化烟酰胺腺嘌呤二核苷酸(NAD+)与NADH (NAD的还原形式)的比值和抗氧化超氧化物歧化酶(SOD)活性显著降低。图2 g),提示膳食胆固醇可诱导肝脏氧化应激。总的来说,这些发现表明,在hfhc喂养的小鼠的非hcc肝组织中形成了NASH和纤维化。
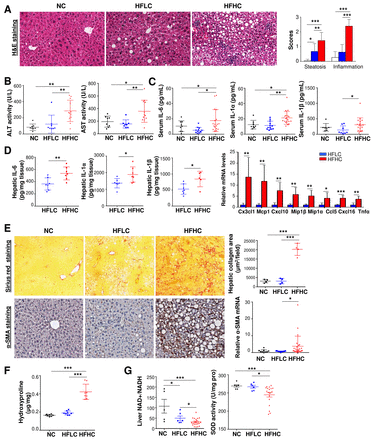
胆固醇引起高脂高脂饲料喂养的小鼠非肝细胞癌肝组织的NASH和纤维化。(A)代表性H&E染色,肝切片组织学评分;(B)血清ALT和AST水平;(C) NC、HFLC和HFHC饲喂14个月小鼠血清IL-6、IL-1α和IL-1β蛋白水平的细胞因子谱分析;(D)用ELISA法测定小鼠肝脏促炎因子IL-6、IL-1α和IL-1β蛋白水平,用RNA测序测定小鼠体内Cx3cl1、Mcp1、Cxcl10、Mip1β、Mip1α、Ccl5、Cxcl16、Tnfα mRNA水平;(E)胶原蛋白沉积,天狼星红染色,免疫组化染色,RT-PCR检测α-SMA蛋白和mRNA水平;(F)饲喂NC、HFLC和HFHC 14个月小鼠肝脏羟脯氨酸含量,(G)肝脏NAD+ / NADH比值和SOD活性。* * * p < 0.05, p < 0.01, * * * p < 0.001。ALT,丙氨酸转氨酶;AST、天冬氨酸转氨酶; α-SMA, alpha-smooth muscle actin; HCC, hepatocellular carcinoma; HFHC, high-fat/high-cholesterol diet; HFLC, high-fat/low-cholesterol diet; H&E, Hematoxylin and eosin; NAD, nicotinamide adenine dinucleotide; NASH, non-alcoholic steatohepatitis; NC, normal chow; RT-PCR, reverse transcription polymerase chain reaction; SOD, superoxide dismutase.
高胆固醇饮食导致小鼠脂肪肝、脂肪性肝炎和纤维化
为了阐明NAFLD在饮食胆固醇形成HCC之前的进展,我们监测了喂食HFHC的小鼠的血清ALT、AST、胆固醇、AFP和肝脏组织学变化。分别于饲粮喂养后3、8、10和12个月收割小鼠(图3一).我们观察到体重和内脏脂肪重量的增加(在线补充图S3A),在3个月和8个月时,与HFLC喂养的小鼠相比,HFHC喂养的小鼠的肝脏重量和肝脏体重比增加(在线补充图S3B).血清ALT和AST升高(在线补充图S3C)与高果糖hhc喂养的小鼠血清胆固醇水平的提高一致(在线补充图S3D).hfhc喂养的小鼠肝脏组织在3个月时显示脂肪变性伴轻度炎症,8个月时显示脂肪性肝炎伴纤维化,10、12和14个月时肝细胞形成,而hflc喂养的小鼠在3、8、10和14个月时仅显示脂肪变性,未出现进一步的肝细胞发展(图3 b而且在线补充图S4A-B).我们还对两种HCC标志物AFP和高尔基蛋白73 (GP73)进行了免疫组化。hfhc喂养的小鼠肝脏组织呈阳性染色,而hflc喂养的小鼠则无阳性染色(图3 b而且在线补充图S4A).肝脏脂肪变性、小叶炎症和肝脏胶原区的评分证实了肝脏组织学在各个疾病阶段的严重程度增加(图3 b).此外,10%(1/10)的hfhc喂养的小鼠在10个月时发生HCC(肿瘤数目=0.1±0.32,最大肿瘤直径=0.55±1.74 mm), 25%(3/12)的小鼠在12个月时发生HCC(肿瘤数目=0.25±0.45,最大肿瘤直径=1.57±3.10 mm), 68%(13/19)的小鼠在14个月时发生HCC (图3 c),表明hfhc喂养的小鼠逐渐发展为脂肪变性、脂肪性肝炎、纤维化和NAFLD-HCC。

hfhc喂养的小鼠依次出现脂肪肝、脂肪性肝炎、纤维化和肝细胞癌。(A) HFHC饲料对C57BL/6小鼠的治疗示意图。小鼠在3、8、10和12月龄时被处死。(B)分别饲喂HFLC 14个月和HFHC 3、8、10、12、14个月小鼠肝脏肿瘤标志物AFP和GP73的代表性大体形态、H&E染色和免疫组化染色;计算脂肪变性和炎症组织学评分和天狼星红染色定量。(C)不同时间点喂食HFHC小鼠最大肿瘤的肿瘤发生率、肿瘤数量和最大肿瘤直径。* * * p < 0.05, p < 0.01, * * * p < 0.001。据法新社,甲胎蛋白;肝癌,肝细胞癌;饲料、高脂肪、高胆固醇的饮食; HFLC, high-fat/low-cholesterol diet; H&E, hematoxylin and eosin; IHC, immunohistochemistry.
膳食胆固醇诱导的肠道菌群失调在NAFLD-HCC的起始和进展中的作用
为了探索肠道菌群在介导膳食胆固醇诱导的NAFLD-HCC中的潜在作用,我们在14个月时对hflc喂养的小鼠和hfhc喂养的小鼠的粪便进行了16S rRNA基因测序。稀疏分析表明,观察到的操作分类单元数量达到饱和(在线补充图S5A-B).采用加权Unifrac主成分分析(PCA),观察hflc和hfhc喂养小鼠在14月龄时肠道菌群组成的差异(p<0.001) (图4一).此外,在14个月时,与单纯脂肪变性的HFLC饮食喂养的小鼠相比,hfhc喂养的HCC小鼠观察到细菌多样性较低,而细菌丰富度增加(图4一).用差异丰度法测定胆固醇相关细菌类群。与hflc喂养的小鼠相比,HFHC喂养的小鼠中几种细菌OTUs的丰度存在差异(图4 b,在线补充表S1-5).主成分和冗余分析还显示,喂食HFHC 3个月(单纯脂肪变性伴轻度炎症)、8个月(脂肪性肝炎伴纤维化)和14个月(肝细胞癌)小鼠的菌群组成明显聚集。图4 c)表明随着NAFLD-HCC进展阶段肠道菌群的组成变化。此外,随着NAFLD-HCC进展,细菌丰富度依次增加(图4 d).特别是,Mucispirillum schaedleri_Otu038, Desulfovibrio_Otu047, Anaerotruncus_Otu107而且Desulfovibrionaceae_Otu073被观察到依次从3个月增加到8个月和14个月的高氢丙烃喂养(图4 d).此外,我们还观察到梭状芽胞杆菌辣子鸡等celatum_Otu070梭状芽胞杆菌,c . ruminantium_Otu059,c . cocelatum_Otu036而且c . methylpentosum_Otu053,和消耗双歧杆菌_otu026, Akkermansia市政phila_otu034,乳酸菌_otu009,拟杆菌_酸化otu032,拟杆菌_otu012, B. uniformis_Otu080而且b . eggerthii_Otu079高胆固醇的人。此外,HFHC-enriched幽门螺杆菌ganmanii_Otu031与没有肿瘤的小鼠相比,hhc喂养的肿瘤小鼠中hhc含量更高,而hhc消耗的Bacteroides_Otu012与未患肿瘤的小鼠相比,高脂高脂小鼠的肿瘤细胞减少(在线补充图S5C).这些建议进一步丰富幽门螺杆菌ganmanii_Otu031和损耗Bacteroides_Otu012可能对肠道菌群在NAFLD-HCC中的作用很重要。此外,与hflc喂养的小鼠相比,hfhc喂养的小鼠肠道菌群相关色氨酸代谢能力降低(图4 e).综上所述,表明高胆固醇饮食诱导的肠道菌群失调和微生物色氨酸代谢受损。
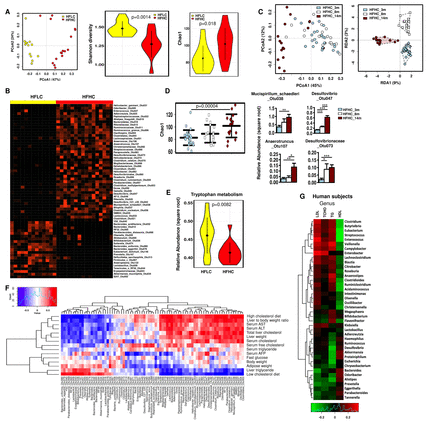
膳食胆固醇诱导的肠道菌群失调。(A) 14月龄HFLC和hfhc喂养小鼠的主成分排序分析(PcoA)、Shannon多样性和肠道菌群丰富度。(B)喂食HFLC或HFHC 14个月小鼠粪便中细菌的热图图。(C)分别饲喂HFHC 3个月、8个月和14个月时肠道菌群的PCoA和冗余分析。(D) HFHC饲喂3、8、14个月时,用chao1指数测定菌群丰富度,依次增加菌群。(E) hflc和hfhc喂养小鼠肠道菌群色氨酸代谢能力。(F)细菌丰度与小鼠表型的相关性。(G) 59例高胆固醇血症患者和39名健康受试者的细菌宏基因组测序与血清总胆固醇、甘油三酯、低密度脂蛋白胆固醇和高密度脂蛋白胆固醇的关系。* * * p < 0.05, p < 0.01, * * * p < 0.001。据法新社,甲胎蛋白; ALT, alanine aminotransferase; AST, aspartate aminotransferase; HDL, high-density lipoprotein; HFHC, high-fat/high-cholesterol diet; HFLC, high-fat/low-cholesterol diet; LDL, low-density lipoprotein; TCHO, total cholesterol; TG, triglyceride.
进行相关分析以确定细菌丰度与小鼠表型的潜在关联。我们观察到,m . schaedleri_Otu038日,Desulfovibrio_Otu047, Anaerotruncus_Otu107, C celatum_Otu070,c . cocelatum_Otu036而且c . methylpentosum_Otu053与hfhc喂养的小鼠粪便样本中富集的图4 f),而双歧杆菌_otu026, B. acidifaciens_Otu032, B. uniformis_Otu080, A. muniphila_otu034而且Lactobacillus_Otu009,与高胆固醇饮食、血清和肝脏胆固醇水平呈负相关(图4 f).这些结果表明,NAFLD-HCC中肠道菌群失调与胆固醇水平相关。
为了证实我们在人类患者动物实验中的发现,我们分析了59例高胆固醇血症患者和39名健康受试者的血清胆固醇和肠道菌群的相关性。这98个被试的特征显示在联机补充表S6.双歧杆菌属而且拟杆菌与血清总胆固醇、低密度脂蛋白(LDL)-胆固醇负相关,与高密度脂蛋白(HDL)-胆固醇正相关(图4 g).这些结果与观察到的hhc喂养的小鼠(在线补充表S1-5),进一步推断肠道菌群参与了胆固醇诱发的紊乱。
膳食胆固醇通过诱导代谢物改变促进NASH-HCC进展
为了揭示与肠道微生物组相关的代谢表型,这可能涉及胆固醇诱导的NAFLD-HCC,我们对hfhc喂养的小鼠和hflc喂养的小鼠的血清进行了代谢谱分析。血清代谢产物根据膳食胆固醇含量存在显著差异(图5一个而且联机补充表S7).胆汁酸生物合成是高胆固醇饮食改变小鼠的一个关键途径(图5 b).初级胆汁酸包括牛磺酸胆酸(TCA),牛磺酸脱氧胆酸(TUDCA),糖胆酸(GCA)和牛磺酸脱氧胆酸(TCDCA)在hfhc喂养的小鼠中是异常值上调的代谢物(图5 c).另一方面,微生物色氨酸代谢的产物3-吲哚丙酸(IPA)18是hhc喂养小鼠的异常值下调代谢物(图5 c).此外,与hflc喂养的小鼠相比,hfhc喂养的小鼠门静脉血清脂多糖(LPS)浓度升高,结肠e -钙粘蛋白丢失(图5 d),表明饮食中的胆固醇会损害肠屏障功能。进行相关分析以确定hhc改变的微生物和代谢物的潜在联系。相关分析一致显示hfhc富集Mucispirillum schaedleri_Otu038与TUDCA、TCDCA、TCA、GCA呈正相关。HFHC-enrichedRoseburia_Otu056而且Helicobacter_ganmanii_Otu031与TUDCA、TCDCA呈正相关。此外,HFHC-depletedAkkermancia_muciniphila_Otu034与TCDCA和TUDCA呈负相关,而hfhc富集Anaerotruncus_Otu107与IPA (图5 e).
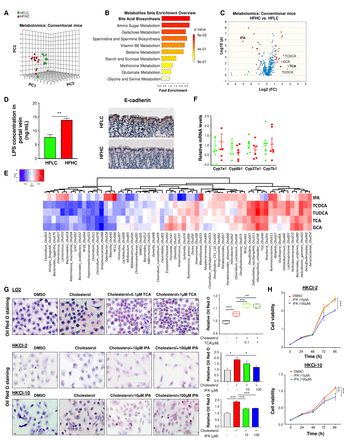
膳食胆固醇诱导的小鼠血清代谢谱的改变。(A)通过主成分排序分析,HFLC和HFHC饲粮组小鼠血清代谢物差异显著。(B) HFHC饲粮小鼠差异富集代谢物的途径分析。(C) HFLC和hhfhc饲粮小鼠血清代谢组学火山图,代谢物异常值显示。(D) HFLC和HFHC饲粮喂养3个月的小鼠门静脉血LPS浓度和14个月HFLC和HFHC饲粮喂养的小鼠结肠组织E-cadherin表达。(E) hfhc改变的微生物与代谢物的相关性分析。(F) HFLC和HFHC饲粮小鼠肝组织中Cyp7a1、Cyp8b1、Cyp27a1和Cyp7b1 mRNA水平。(G) TCA加重人LO2细胞系胆固醇诱导的甘油三酯积累,IPA抑制NASH-HCC细胞系、HKCI-2和HKCI-10细胞系胆固醇诱导的甘油三酯积累。胆固醇,200μg / mL;(H) IPA抑制NASH-HCC细胞系细胞增殖。 *p<0.05, **p<0.01, ***p<0.001. DMSO, dimethyl sulfoxide; GCA, glycocholic acid; HCC, hepatocellular carcinoma; HFHC, high-fat/high-cholesterol diet; HFLC, high-fat/low-cholesterol diet; IPA, 3-indolepropionic acid; NASH, non-alcoholic steatohepatitis; LPS, lipopolysaccharides; TCA, taurocholic acid; TCDCA, taurochenodeoxycholic acid; TUDCA, tauroursodeoxycholic acid.
进一步的实验发现,高胆固醇并不会改变肝脏中细胞色素P450 (Cyp)7a1、Cyp8b1、Cyp27a1和Cyp7b1等胆汁酸合成酶mRNA的表达(图5 f).体外实验表明,TCA可加重人正常永生化肝细胞LO2中胆固醇诱导的甘油三酯积累(图5克),而IPA则抑制胆固醇引起的脂质积聚(图5克)和细胞增殖(图5 h)在NASH-HCC细胞系中,HKCI-2和HKCI-10。这些结果表明,胆固醇通过调节宿主血清代谢物,以及至少部分通过增加TCA和减少IPA,促进了NASH-HCC的进展。
高胆固醇调节的肠道菌群促进无菌小鼠脂肪性肝炎和肝细胞增殖
我们通过粪便菌群移植(FMT)评估胆固醇调节菌群在NAFLD-HCC中的作用。将NC、hflc和hfhc喂养的小鼠(14月龄)的粪便灌胃给NC饲粮下的无菌小鼠(G-NC、G-HFLC和G-HFHC)。图6).G-HFHC小鼠的肝重比显著高于G-NC小鼠(在线补充图S6A).此外,甘油三酯试验和油红O染色(图6 b)和硫代巴比妥酸反应物质测定过氧化作用(图6 b)显著增加,伴随肝脏组织损伤(H&E染色)(图6 b)在FMT后8、10和14个月G-HFHC小鼠中。肝脏细胞因子和包括IL-6在内的趋化因子的增强证实了炎症的增加(图6 c), Fos、Ccl12、Cxcr1、Ccl1、Myd88、Il-1β、Cxcl10和C3ar1 (在线补充表S8)和增强CD45+淋巴细胞积累(在线补充图S6B)流式细胞术检测G-HFHC小鼠与G-HFLC小鼠的差异。

高胆固醇调节菌群促进无菌小鼠肝细胞增殖。(A)将NC、hflc和hfhc喂养的小鼠(14个月)的粪便移植到NC喂养的无菌小鼠(G-NC、G-HFLC和G-HFHC)。G-NC组、G-HFLC组和GHFHC组灌胃粪便的无菌小鼠肝脏的大体形态学、组织学检查和Ki-67染色显示。(B) G-NC组、G-HFLC组和G-HFHC组无菌受体小鼠8、10和14月龄时的肝脏甘油三酯含量、脂质过氧化和肝脏组织学。(C) G-NC、G-HFLC或G-HFHC组14个月时肝脏IL-6蛋白水平和(D)肝切片Ki-67染色。(E1) G-NC, G-HFLC或G-HFHC小鼠肝组织中的小鼠癌症通路Finder PCR阵列。(E2)检测CDC20蛋白水平。(F) G-NC、G-HFLC或G-HFHC小鼠肠道菌群线性判别分析(LDA)评分的主成分排序分析和直方图。(G) G- hflc和G- hfhc小鼠血清代谢组学分析。* * * p < 0.05, p < 0.01, * * * p < 0.001。 HFHC, high-fat/high-cholesterol diet; HFLC, high-fat/low-cholesterol diet; NC, normal chow; PCA, principal component analysis.
G-HFHC小鼠14月龄时肝细胞增殖增加(p<0.05) (图6 d),但在FMT后8个月和10个月没有。与G-NC和G-HFLC小鼠相比,G-HFHC小鼠肝组织中细胞增殖(Cdc20)、血管生成(Pgf)、侵袭/转移(Serpinb2、Snai3)等致癌通路相关基因表达上调,凋亡相关基因(Fasl和Lpl)表达下调。图6 e1).Western blot验证CDC20表达上调(图6 e2).G-HFHC小鼠在14月龄时观察到一个肝结节,但在8月龄和10月龄时未观察到(图6).组织学检查证实结节呈非典型增生,伴球状及细胞增殖增加(图6).我们评估了受体无菌小鼠和相应供体常规小鼠的肠道菌群组成。我们发现,接受无菌小鼠的肠道菌群与供体饮食有显著差异(图6 f).各组小鼠在FMT后的微生物生态系统随时间的推移保持稳定,β-多样性和组成分析(在线补充图S7A-D).此外,对无菌小鼠的血清代谢物的分析显示,与G-HFLC小鼠相比,G-HFLC小鼠的IPA下降,与常规小鼠饲喂HFHC (图6克).综上所述,这些数据表明,饮食中胆固醇调节的微生物群通过诱导代谢物改变促进NAFLD和肝细胞增殖,从而促进胆固醇诱导的NAFLD - hcc的形成。
抗胆固醇治疗完全防止NAFLD-HCC在hfhc喂养的小鼠中形成
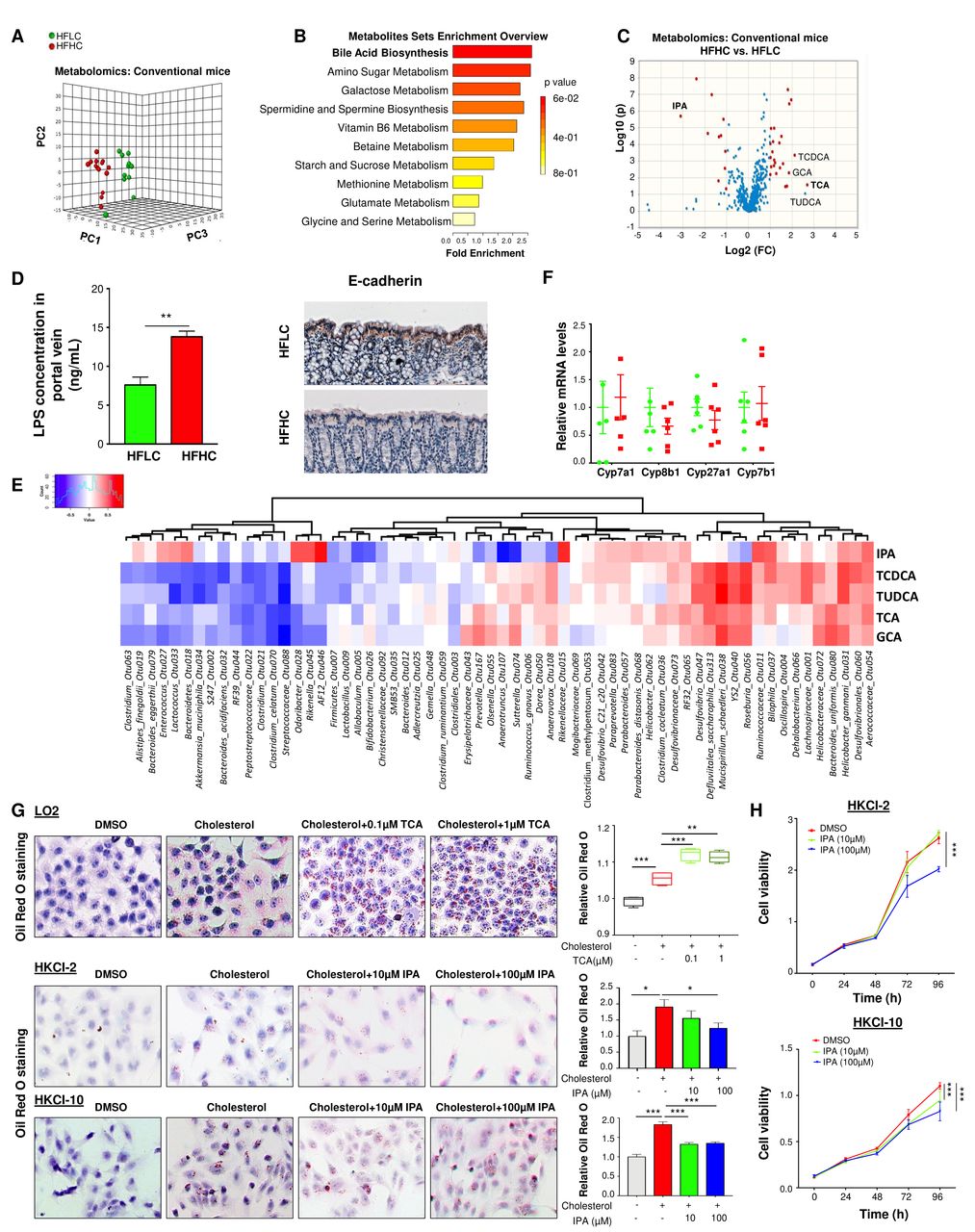
由于膳食胆固醇驱动NAFLD - HCC的进展,我们评估了抗胆固醇药物是否可以抑制NAFLD及其进展为HCC。用降胆固醇药物阿托伐他汀(20 mg/kg)喂喂喂高脂醚7个月的小鼠,并继续喂喂高脂醚和阿托伐他汀7个月(图7).试验结束时(14个月),阿托伐他汀完全阻止了HFHC饮食诱导的NAFLD-HCC的形成,并改善了NASH的严重程度(图7 b).与此同时,血清胆固醇、肝游离胆固醇、血清甲胎蛋白(图7 c)、ALT、降低血清促炎细胞因子(IL-6、IL-1α、IL-1β、MCP-1、MIP-1α和MIP-1β)和氧化应激(增加NAD+ / NADH比值和SOD活性)(图7 d).阿托伐他汀通过显著降低肝脏胶原沉积和羟脯氨酸含量(图7 e).采用16S rRNA基因测序法对阿托伐他汀治疗组(HFHC+At)灌喂HFHC小鼠的粪便进行菌群分析,并与NC、HFLC和HFHC灌喂小鼠的粪便进行比较。经抗胆固醇阿托伐他汀治疗后,小鼠体内增加的细菌丰富度显著恢复(图7 f).此外,在hfhc喂养的小鼠中,调节异常的OTUs (图4B和D),丰富的Mucispirillum schaedleri_Otu038, Desulfovibrio_Otu047, Anaerotruncus_Otu107而且Desulfovibrionaceae_Otu073经抗胆固醇治疗后(图7 f).为了研究菌群在抗胆固醇药物预防的NAFLD-HCC中的直接作用,我们将HFHC+At小鼠的粪便灌胃给无菌小鼠(G-HFHCAt, n=10)。灌胃后14个月,G-HFHC组小鼠肝脏组织改善,肝脏甘油三酯和脂质过氧化降低(图7 g).用阿托伐他汀治疗HFHC小鼠,观察到IPA的诱导和TCA的降低。联机补充图S8).这些结果进一步表明,肠道菌群在介导胆固醇诱导的NAFLD-HCC中发挥了积极作用。

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
胆固醇抑制阻断了hfhc喂养的小鼠NAFLD-HCC的进展。(A)阿托伐他汀对HFHC饲粮喂养的C57BL/6小鼠的治疗示意图。(B) h&e染色的hfhc喂养的小鼠,在阿托伐他汀治疗或未治疗的情况下,计算肝脏代表性的毛形态和显微镜特征,脂肪变性、炎症和胶原蛋白的组织学评分。(C)阿托伐他汀治疗前后hfhc喂养小鼠的血清胆固醇水平、肝脏游离胆固醇水平、血清AFP水平、(D)血清ALT水平、肝脏NAD+ / NADH比值、肝脏SOD活性、血清IL-6、IL-1α、IL-1β、MCP-1、MIP-1α、MCP-1β蛋白水平、(E)胶原沉积和羟脯氨酸含量。(F)饲喂NC、HFLC和HFHC(加或不加阿托伐他汀治疗)小鼠粪便中细菌丰富度和细菌热图图。(G)灌胃HFHCAt粪便的无菌小鼠(G-HFHCAt)的大体形态、组织学检查、肝脏甘油三酯和脂质过氧化情况。(H)胆固醇诱导的NAFLD-HCC发生机制示意图。* * * p < 0.05, p < 0.01, * * * p < 0.001。据法新社,甲胎蛋白;At,阿托伐他汀,ALT,丙氨酸氨基转移酶; HCC, hepatocellular carcinoma; HFHC, high-fat/high-cholesterol diet; IPA, 3-indolepropionic acid; NAD, nicotinamide adenine dinucleotide; NAFLD, non-alcoholic fatty liver disease; ROS, reactive oxygen species; SOD, superoxide dismutase; TCA, taurocholic acid.
讨论
尽管胆固醇是NASH中已知的细胞毒性脂质,5 8关于胆固醇在NASH HCC发展中的作用和重要机制的信息有限。在这项研究中,我们首次证明了长期高胆固醇饮食喂养会导致小鼠自发性NAFLD-HCC的发展。高脂饮食可诱发肥胖、胰岛素抵抗、葡萄糖不耐受和脂肪变性。然而,脂肪性肝炎向纤维化和肝细胞癌的进展主要与高饮食胆固醇有关。移植了高胆固醇饮食喂养小鼠粪便菌群的无菌小鼠在脂质积累、炎症和细胞增殖方面基本复制供体小鼠,这表明肠道菌群失调有助于饮食胆固醇诱导的NASH和NAFLD-HCC的潜在发展。
我们调查了高饮食胆固醇导致NAFLD发生HCC的重要机制。我们发现高胆固醇饮食喂养的小鼠NAFLD-HCC中ROS积累增强。累积的ROS是一种毒性介质,可诱导炎症反应、胰岛素抵抗和氧化损伤。19事实上,我们的分析显示,膳食胆固醇诱导ROS和促炎细胞因子,从而促进NASH和HCC的发展。这和我们的发现一致,沃尔夫等研究表明,长期饲喂胆碱缺乏的高脂饮食可诱导小鼠肝脏ROS、肝内CD8+ T细胞、NKT细胞及其分泌的炎症细胞因子促进NASH和HCC的转变。20.
越来越多的证据表明,人类肠道菌群可以影响包括肝癌在内的各种病理条件。21因此,我们探索了肠道菌群在介导膳食胆固醇诱导的NAFLD-HCC中的潜在参与。随着细菌丰富度的依次增加,M. schaedleri_Otu038, Desulfovibrio_Otu047, Anaerotruncus_Otu107而且Desulfovibrionaceae_Otu073随着NAFLD-HCC的进展而增加。这与报告一致m . schaedleri而且Desulfovibrionaceae在小鼠模型中与肥胖、代谢综合征和炎症密切相关。22 - 24Desulfovibrionaceae而且脱磷孤菌属在猪NASH模型中被描述为丰富的。25丰富Anaerotruncus在MCD饮食诱导的实验性NASH小鼠模型中也有报道。26此外,我们还观察到几个梭状芽胞杆菌OTUs富含高胆固醇饮食,与高血清胆固醇呈正相关。以前的报告显示梭状芽胞杆菌在纳什的病人,27支持我们在本研究中的观察。此外,OTUs的损耗Akkermansia、乳酸菌、双歧杆菌而且拟杆菌在NAFLD-HCC小鼠中得到证实。这些辣子鸡包括A. muniphila_otu034,乳酸菌_otu009,双歧杆菌_otu026, B. uniformis_Otu080而且b . acidifaciens_Otu032与高血清胆固醇一致降低。双歧杆菌属而且拟杆菌在本研究中进一步显示与人类高胆固醇血症患者的血清总胆固醇呈负相关。损耗的双歧杆菌属而且拟杆菌已经在人类NASH患者中得到证实。27 28有证据表明双歧杆菌属通过调节肠道菌群具有降低胆固醇的活性。此外,b . acidifaciens是一种潜在的益生菌,因为它具有预防肥胖和提高小鼠胰岛素敏感性的能力。29的应变b .均匀化,即B. uniformis CECT 7771据报道可改善小鼠高脂饮食引起的代谢和免疫功能障碍。30.菌株的乳酸菌也被报道可以预防NASH31并以与他汀类药物治疗小鼠模型相似的模式对肝脏胆固醇代谢产生有益影响。32此外,减少答:muciniphila据报道,NAFLD通过稀释肠道黏液层,从而损害肠道的通透性屏障。33这些结果支持了我们在这项研究中的观察,并进一步表明抑制保护性肠道微生物可能是胆固醇在肝脏中发挥其致癌作用的一种手段。我们的结果为肠道菌群失调在胆固醇诱导的NAFLD-HCC进展中的潜在作用提供了新的见解。
除了肠道菌群失调外,共生菌产生的小分子代谢物可能是NAFLD发病的原因之一。13肠道微生物相关代谢物,包括短链脂肪酸、氨基酸分解代谢产物和胆胆酸,都可以使同源受体感到痛苦和拮抗,从而减少或加剧肝脏脂肪变性和炎症。34最近,一种新的肠道菌群代谢产物N,N,N-三甲基-5-氨基戊酸(TMAVA)被发现在人肝脂肪变性的血浆中升高,并加剧小鼠脂肪肝的进展。35因此,我们试图解开可能介导高饮食胆固醇对NAFLD-HCC促进作用的代谢物。代谢组学分析显示,高胆固醇饮食会损害胆汁酸代谢。包括TCA、GCA、TCDCA和TUDCA在内的主要胆汁酸在hfhc喂养的小鼠中富集,与先前报道的与健康受试者相比的人类NASH患者血浆TCA、GCA和TCDCA较高一致。36破坏胆汁酸稳态会改变肝脏的代谢稳态,并可能导致肝脏炎症和代谢性疾病,包括糖尿病和NAFLD。37 38TCA、GCA、TCDCA和TUDCA是连接肠道和肝脏的关键信号分子,影响肝脏脂质积累和炎症。39双歧杆菌属而且拟杆菌是参与胆汁酸代谢的肠道菌群的主要细菌属。40它们可以通过胆汁酸水解酶将牛磺酸结合和甘氨酸结合的胆汁酸解耦为未结合的游离形式,并将未结合的一级胆汁酸转化为次生胆汁酸。40因此,减少双歧杆菌属而且拟杆菌可能解释了hfhc喂养的小鼠体内牛磺酸结合胆汁酸的积累。在这项研究中,TCA在体外加剧了胆固醇诱导的甘油三酯的积累,这进一步支持了它和其他丰富胆汁酸在胆固醇诱导的NAFLD-HCC发病机制中的潜在作用。此外,IPA,完全由共生肠道微生物产生,41 42在hfhc喂养的常规小鼠和移植了hfhc喂养小鼠的粪便的无菌小鼠体内都是耗尽的。IPA是一种特殊的代谢物,其产生完全依赖于肠道菌群对色氨酸的代谢18具有抗炎和保护肠道屏障完整性的特点。42我们发现微生物色氨酸代谢显著降低。体外功能分析显示IPA可抑制NASH-HCC细胞增殖和抑制脂质积累。总的来说,我们的研究结果表明,胆固醇可损害胆汁酸代谢和微生物色氨酸代谢,导致血清TCA增强,IPA降低,从而促进NAFLD-HCC的发展(图7 h).
通过粪便移植的肠道菌群转移已被用于证明肠道菌群在代谢性疾病(包括肥胖和NAFLD)中的积极作用。27 43因此,我们试图确定肠道菌群是否可以在膳食胆固醇诱导的肝脂肪性肝炎和肿瘤发生中发挥直接的诱因作用。与对照组相比,胆固醇调节菌群促进无菌小鼠脂肪性肝炎和肝细胞增殖/发育不良。在无菌小鼠中,促炎因子和IPA降低与微生物群诱导的肝脂肪性肝炎和肿瘤发生相关,与常规小鼠模型的观察结果一致。除了肝脂积聚、炎症、细胞增殖和异型增生外,移植高胆固醇饮食喂养小鼠粪便菌群的无菌小鼠在14个月后未观察到HCC。这表明,膳食胆固醇在诱导NAFLD-HCC的形成中起着重要作用,膳食胆固醇改变肠道菌群有助于疾病的进展。
如果胆固醇在NASH和HCC的发病机制中起着关键作用,那么确定其功能阻断可以改善脂肪性肝炎的严重程度和NAFLD-HCC的发展就很重要了。为了验证这一点,我们使用了阿托伐他汀,一种广泛用于治疗高胆固醇血症的3-羟基-3-甲基戊二酰辅酶a (HMG-CoA)还原酶抑制剂,来抑制hfhc喂养小鼠的胆固醇生物合成。我们发现在阿托伐他汀治疗的小鼠中没有发现HCC,脂肪性肝炎和纤维化的严重程度大大减弱。此外,细菌丰富,Mucispirillum,脱磷孤菌属,Anaerotruncus而且Desulfovibrionaceae随着胆固醇诱导的NAFLD-HCC进展增加,阿托伐他汀治疗逆转。本研究显示,恢复肠道菌群多样性可能是阿托伐他汀对NAFLD-HCC产生保护作用的机制之一。此外,与用只喂食HFHC的小鼠粪便灌胃的小鼠不同,用阿托伐他汀治疗的HFHC喂养的小鼠的粪便不促进无菌受体小鼠的肝细胞增殖。这些观察结果进一步强调了阿托伐他汀在预防胆固醇诱导的NAFLD-HCC发展方面的潜在用途。
总之,本研究首次表明,长期的高饮食胆固醇通过调节肠道菌群诱导雄性小鼠NAFLD-HCC的自发和进行性发展。胆固醇通过肠道菌群的改变诱导TCA升高,IPA降低,从而促进肝脏中脂质积累、细胞增殖,导致NAFLD-HCC的发展(图7 h).抗胆固醇治疗完全消除了饮食胆固醇诱导的NAFLD-HCC的形成。本研究强调,胆固醇抑制和操纵肠道菌群及其相关代谢物可能是预防NAFLD-HCC的有效策略。
参考文献
脚注
XZ和OOC是共同第一作者。
贡献者XZ和OOC参与研究设计,进行实验并撰写论文;EC、KF、HCHL和Y-XW进行实验;AWHC确定组织学;HW提供无菌动物;XY和JJYS对稿件进行了批改;JY设计、指导并撰写了论文。
资金本项目获广东省自然科学基金(2018B030312009)、研资局协作研究基金(C4041-17GF, C7026-18G, C7065-18G)、香港研资局主题研究计划(t12 -703/ 19r)、香港中文大学研究直接补助金、香港中文大学校长自由支配基金资助。
相互竞争的利益没有宣布。
病人同意发表不是必需的。
伦理批准所有动物研究均按照香港中文大学和第三军医大学动物实验伦理委员会批准的准则进行。
来源和同行评审不是委托;外部同行评议。
数据可用性声明所有与研究相关的数据都包含在文章中或作为补充信息上传。