条文本

摘要
客观的虽然在COVID-19患者的粪便中检测到严重急性呼吸综合征冠状病毒2型(SARS-CoV-2) RNA,但疾病过程中该病毒在胃肠道中的活性和传染性在很大程度上尚不清楚。我们研究了SARS-CoV-2的时间转录活性及其与COVID-19患者粪便微生物组纵向变化的关系。
设计我们对15名COVID-19住院患者的系列粪便病毒提取进行了RNA鸟枪宏基因组测序。对SARS-CoV-2基因组测序覆盖率进行了量化。我们评估了粪便微生物组组成和微生物组功能与粪便SARS-CoV-2传染性特征的相关性。
结果经病毒RNA宏基因组测序,15例新冠肺炎患者中有7例(46.7%)粪便检测出SARS-CoV-2阳性。即使没有胃肠道表现,所有7名患者在其粪便病毒宏基因组中SARS-CoV-2基因组3 '端(p=0.0261)和5 '端(p=0.0094)的覆盖率和密度都显著更高。在从呼吸道样本中清除SARS-CoV-2后6天,三名患者的粪便病毒宏基因组继续显示活跃的病毒感染特征(3 ' vs 5 '端覆盖率更高)。具有高传染性特征的粪便样本具有较高的细菌种类丰度Collinsella aerofaciens,Collinsella tanakaei,链球菌对象,摩根氏菌属morganii具有较高的核苷酸从头生物合成、氨基酸生物合成和糖酵解功能能力,而具有低至无传染性特征的粪便样本具有较高的短链脂肪酸生成细菌丰度,Parabacteroides merdae,拟杆菌stercoris,Alistipes onderdonkii而且Lachnospiraceae细菌1_1_57FAA.
结论这项初步研究提供了证据,即使在没有胃肠道表现和从SARS-CoV-2呼吸道感染中恢复后,也存在活跃和长期的“静止”胃肠道感染。SARS-CoV-2 GI感染活动性患者的肠道菌群特点是机会致病菌丰富,有益细菌减少,核苷酸和氨基酸生物合成功能和碳水化合物代谢功能增强。
- 肠道炎症
- 传染性疾病
- 诊断病毒学
这是一篇开放获取的文章,根据创作共用署名非商业(CC BY-NC 4.0)许可证发布,该许可证允许其他人以非商业方式分发、混音、改编、在此基础上进行构建,并以不同的条款许可其衍生作品,前提是正确引用原始作品,给予适当的荣誉,任何更改都已注明,并且使用是非商业性的。看到的:http://creativecommons.org/licenses/by-nc/4.0/.
数据来自Altmetric.com
本研究的意义
关于这个问题我们已经知道了什么?
相当一部分COVID-19患者存在胃肠道症状。
在一些COVID-19患者的呼吸道样本中,即使病毒RNA呈阴性,但仍在其粪便样本中检测到严重急性呼吸综合征冠状病毒2型(SARS-CoV-2) RNA。
对SARS-CoV-2感染细胞模型的体外转录分析表明,SARS-CoV-2基因组3 '端比5 '端被高度覆盖,表明病毒复制和感染活跃。
新的发现是什么?
我们首次在一个亚组(47%)的COVID-19患者中发现了活跃肠道病毒感染的特征,即使在没有胃肠道症状的情况下,这表明SARS-CoV-2的“静止”胃肠道感染。
即使在呼吸道清除SARS-CoV-2后,病毒感染和复制的转录活性仍然存在于肠道中。
具有SARS-CoV-2高传染性特征的粪便样本含有更多的机会性病原体,Collinsella aerofaciens,Collinsella tanakaei,链球菌对象,摩根氏菌属morganii核苷酸和氨基酸生物合成以及碳水化合物代谢(糖酵解)的能力增强,而具有低至无SARS-CoV-2传染性特征的粪便样本具有更高丰度的短链脂肪酸生成细菌,Parabacteroides merdae,拟杆菌stercoris,Alistipes onderdonkii而且Lachnospiraceae细菌1_1_57FAA.
本研究的意义
在可预见的未来,它会对临床实践产生怎样的影响?
COVID-19患者的肠道中SARS-CoV-2活性活跃且持续时间长,即使在没有胃肠道表现和康复后,也突出了长期冠状病毒和健康监测的重要性,以及潜在的粪-口病毒传播的威胁。
应该探索包括消除肠道SARS-CoV-2活性和调节肠道微生物组组成和功能在内的治疗方法。
简介
COVID-19是一种由新型冠状病毒(SARS-CoV-2)引起的急性呼吸道疾病,其特征是病毒在上呼吸道活跃复制。1来自武汉的早期报告显示,2%-10%的COVID-19患者有腹泻等胃肠道症状,但最近的一项荟萃分析报告称,高达20%的患者有胃肠道症状。2 - 5此外,在伴有腹泻的COVID-19患者中,粪便钙保护蛋白(肠道炎症反应的指标)升高。6这些证据表明,消化道可能是COVID-19消化道表现为SARS-CoV-2感染的肺外部位。然而,SARS-CoV-2在COVID-19患者病程期间和疾病消退后胃肠道中的病毒活性和传染性在很大程度上尚不清楚。
基于RT-PCR,在相当一部分COVID-19患者的肛拭子和粪便样本中检测到SARS-CoV-2 RNA。1 7此外,通过电镜观察新冠肺炎患者粪便中SARS-CoV-2病毒颗粒。8在一部分COVID-19患者的呼吸道标本呈病毒阴性后,粪便中SARS-CoV-2 RNA的存在可以持续更长时间7 9.然而,对于呼吸道清除SARS-CoV-2后粪便呈阳性的COVID-19患者,其肠道中SARS-CoV-2病毒的活性尚不清楚。目前,SARS-CoV-2在肠道中的病毒活性主要是根据肠道类器官和哺乳动物细胞模型的间接观察结果推断出来的:(1)SARS-CoV-2受体ACE2在肠道肠上皮细胞和结肠腺细胞中高度表达10 11(2) SARS-CoV-2感染人类和蝙蝠肠道类器官的肠细胞谱系细胞。12日13迄今为止,缺乏人体肠道中具有复制能力和感染能力的SARS-CoV-2病毒的数据。更深入地了解SARS-CoV-2在人类肠道中的生命周期和致病性是一个尚未满足的迫切需求。
在这项初步观察性研究中,我们假设SARS-CoV-2在COVID-19患者的肠道中具有活性,因此描绘了COVID-19住院患者在病程期间和疾病清除后肠道中SARS-CoV-2的转录活性和传染性。我们前瞻性地纳入了2020年2月16日至2020年3月2日在中国香港收治的15名COVID-19住院患者,随访从住院到出院。提取粪便病毒RNA,然后进行鸟枪宏基因组测序和分析,以研究SARS-CoV-2的转录活性。
方法
研究对象与设计
这项前瞻性研究涉及15例因实验室确诊的SARS-CoV-2感染而住院的COVID-19患者(表1).经当地医院和中国香港公共卫生化验所连续两次针对RdRp基因不同区域的RT-PCR检测,确诊为SARS-CoV-2感染。所有COVID-19患者均于2020年2月5日至2020年3月17日期间入住香港威尔士亲王医院或基督教联合医院。他们被随访至出院或至2020年4月4日。所有患者均知情同意参与本研究,并同意发表研究结果。数据包括人口学、流行病学、临床及化验结果,均取自香港医院管理局临床管理系统的电子病历。连续采集COVID-19患者粪便样本,直至出院。
粪便病毒RNA提取和鸟枪宏基因组测序
根据制造商说明,使用TaKaRa MiniBEST病毒RNA/DNA提取试剂盒(TaKaRa,日本)从粪便样本中提取总病毒核酸。提取的病毒总核酸经RNA Clean & Concentrator Kits (Zymo Research, California, USA)纯化得到病毒RNA。通过Qubit 2.0、琼脂糖凝胶电泳和Agilent 2100测试进行质量控制程序后,使用KAPA RNA HyperPrep Kit (Illumina, USA)制备合格的RNA文库。文库制剂在Illumina NextSeq 550平台上测序(配对端150 bp)。
使用Trimmomatic V.0.36对原始序列读取进行过滤和质量调整14如下:1)削减低质量基数(质量分数<20);2)删除小于50 bp的读数;3)跟踪并切断测序适配器。使用默认参数的Kneaddata V.0.5.1(参考数据库:GRChg38)过滤污染的人类读取。
宏基因组测序新冠病毒基因组覆盖度和密度
截至2020年4月20日,从美国国家生物技术信息中心的SAS-CoV-2基因组库下载了1666个完整基因组。通过BBMap V.38.81将清洁的粪便病毒宏基因组读数与定制的SARS-CoV-2参考基因组进行查询。15人工检查了映射的reads,特别是那些映射到SARS-CoV-2基因组3 '端的reads,并丢弃了以Poly A结尾的reads。
将SARS-CoV-2基因组的3 '端定义为从SARS-CoV-2基因组的25 000个核苷酸碱基开始到全长端(~29 9000个核苷酸碱基)的基因组区域,将其余的基因组区域(0-25 000个核苷酸碱基)定义为SARS-CoV-2基因组的5 '端。测序覆盖率被定义为映射到给定基因组区域的霰弹枪测序的数量(每100个SARS-CoV-2基因组核苷酸碱基)。测序密度定义为SARS-CoV-2基因组特定基因组区域测序位点的频率(每500个核苷酸碱基中SARS-CoV-2宏基因组命中数)。
粪便DNA提取,宏基因组测序和细菌微生物组分类和功能分析
用1ml ddH预洗约0.1 g粪便样本2O,以13 000× g离心1 min成粒。随后,按照制造商的说明,使用Maxwell RSC PureFood GMO和认证试剂盒(Promega, Madison, Wisconsin, USA)从颗粒中提取粪便DNA。简单地说,将粪便颗粒加入1 mL十六烷基三甲基溴化铵(CTAB)缓冲液,漩涡30 s,然后在95°C加热样品5 min。之后,用珠粒以最大速度彻底涡旋样品15分钟。然后在样品中加入40µL蛋白酶K和20µL RNase A,在70°C下孵育10 min。13 000× g离心5 min得到上清液,加入Maxwell RSC机进行DNA提取。提取的DNA采用Nextera DNA Flex Library Preparation kit (Illumina)进行DNA文库构建,通过末端修复、尾部加A、纯化、PCR扩增等步骤完成。文库随后在香港中文大学微生物研究中心的Illumina NextSeq 550测序仪(150 bp对端)上测序。人类reads在粪便宏基因组中的比例是通过将reads与参考基因组(human release 32, GRCh38)对齐而产生的。p13,从Gencode下载)由领结2。16
使用Trimmomatic V.0.36对原始序列读取进行过滤和质量调整14(1)削减低质量基数(质量分数<20);(2)去除小于50 bp的读数;3)去除测序适配器。使用默认参数的Kneaddata(参考数据库:GRCh38.p13)过滤污染人体读取。利用MetaPhlAn2 (V 2.9)将reads映射到分支特异性标记,对粪便细菌群落进行分类分析。17使用HUMAnN2.0对粪便细菌群落进行功能分析。18通过LefSE在对应于各自COVID-19患者的所有时间点粪便中鉴定出SARS-CoV-2传染性高和SARS-CoV-2传染性低或无的粪便之间的差异细菌种类。只绘制线性判别分析(LDA)效应量>2和错误发现率(FDR)校正p值<0.05的种。高传染性定义为粪便病毒RNA宏基因组中SARS-CoV-2基因组的3 ' vs 5 '端覆盖率较高。低传染性定义为粪便病毒RNA宏基因组中SARS-CoV-2基因组的3 '和5 '端覆盖相似或没有覆盖。
结果
COVID-19患者
招募了15名实验室确诊的SARS-CoV-2感染的COVID-19住院患者并进行了随访(图1,表1).15例患者中,7例为男性。COVID-19住院患者的中位年龄为55岁(IQR: 44-67)。11例患者为中重度COVID-19患者,2例患者病情危重,入住重症监护病房。所有患者均有呼吸道症状,但只有1例患者(患者5)在发病时同时出现腹泻的胃肠道症状。所有患者在住院期间均无胃肠道症状。从每位患者收集连续的粪便样本,直到患者出院时连续两个样本的鼻咽拭子或喉咙拭子检测呈阴性。中位住院时间为21±2.4天(平均±SE)。

15例COVID-19住院患者的患者症状发作、住院、咽喉(鼻咽)拭子清除严重急性呼吸综合征冠状病毒2 (SARS-CoV-2)和出院的时间轴。
COVID-19患者SARS-CoV-2短暂粪便病毒活性描述
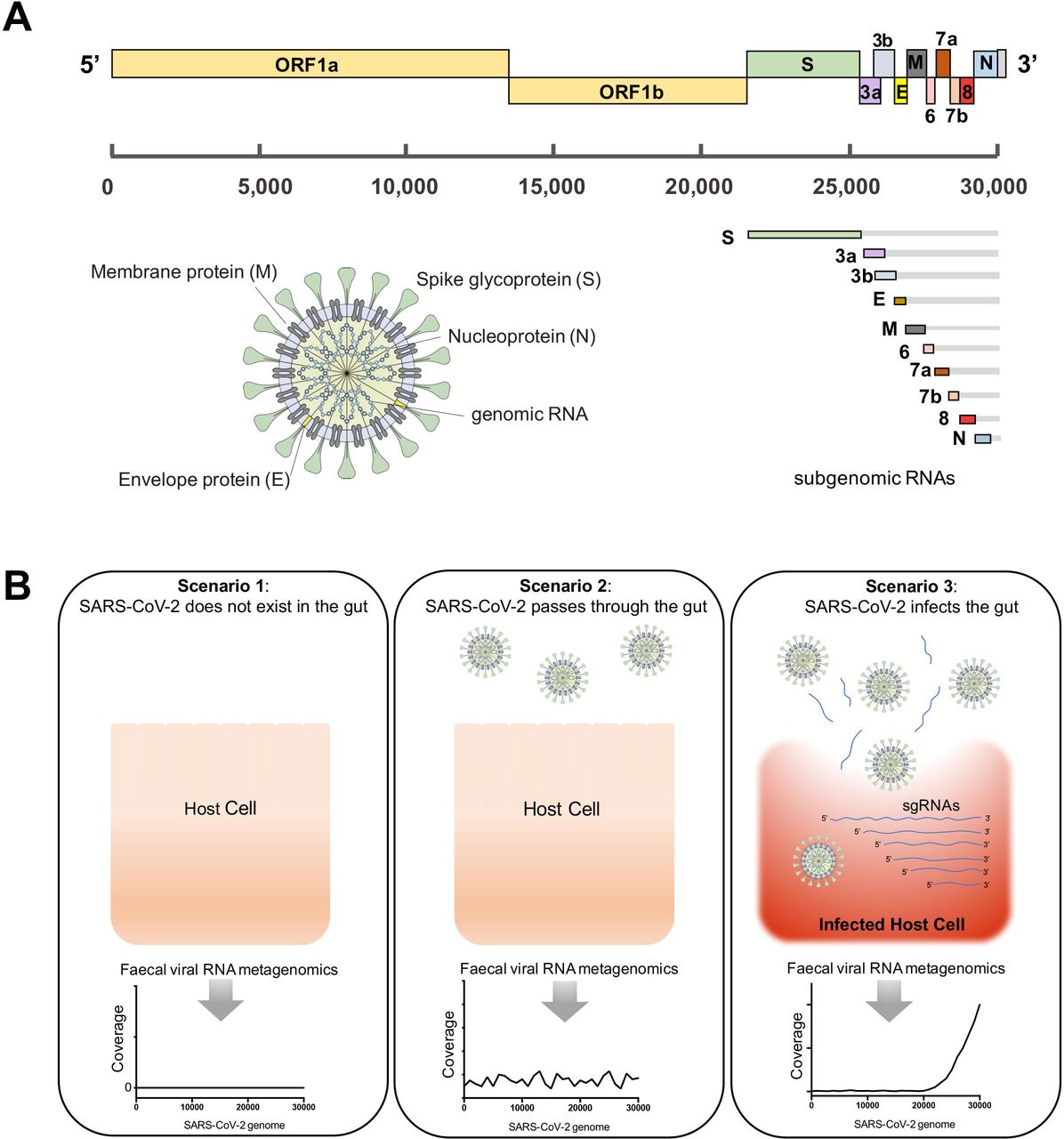
SARS-CoV-2的基因组大小约为29.9 kb核苷酸(图2一个).19日20在进入宿主细胞后,作为mRNA的全长基因组RNA被翻译成ORF1a和ORF1b (SARS-CoV-2基因组的5 '端)。除了全基因组RNA外,还产生了编码结构蛋白S、E、M和N的9种主要亚基因组RNA (sgRNAs) (SARS-CoV-2基因组的3 '端)。20.最近的一项研究表明,通过高通量转录组测序,在感染细胞模型(Vero细胞)中,SARS-CoV-2基因组3 '端比5 '端测序覆盖率高得多。20.这种3 '端sgRNAs的高转录表达是SARS-CoV-2在宿主细胞中活跃复制和感染的标志。

严重急性呼吸综合征冠状病毒2型(SARS-CoV-2)粪便病毒RNA宏基因组谱与肠道传染性的假设情景(A) SARS-CoV-2病毒全长基因组、转录亚基因组rna (sgRNA)和病毒粒子结构示意图。(B)假设COVID-19患者肠道中存在SARS-CoV-2并具有传染性,以及通过粪便病毒RNA气象基因组学测序检测SARS-CoV-2病毒的三种情景。如果SARS-CoV-2病毒感染了肠道内的宿主细胞,其基因组和sgrna应在细胞溶解时高表达并释放到肠腔中,其中SARS-CoV-2基因组的3 '端应被粪便病毒RNA宏基因组测序高度覆盖。
基于这些复制型SARS-CoV-2的转录组模式,我们假设了人类肠道中存在SARS-CoV-2病毒并通过粪便病毒RNA宏基因组学(转录组学)进行检测的三种可能场景:(i)场景1:如果SARS-CoV-2不存在于肠道中,则无法通过宏基因组学测序检测到其RNA基因组;(ii)场景2:如果游离的SARS-CoV-2病毒粒子通过肠道而不感染宿主细胞,则整个病毒RNA基因组应该同样被宏基因组学reads覆盖;(iii)场景3:如果SARS-CoV-2病毒存在于肠道中并感染宿主细胞,通过宏基因组学分析,病毒RNA基因组的3 '端区域应该比5 '端基因组区域覆盖更广(图2 b).
为了描述SARS-CoV-2在COVID-19患者肠道中的病毒转录活性,我们对按时间顺序排列的粪便样本进行了粪便病毒RNA鸟枪宏基因组测序。因此,我们从15例COVID-19患者的连续粪便病毒RNA制剂中平均获得18 773 883±420 617(平均±SE)读数。通过宏基因组测序,7名患者在基线时(住院后第一次粪便采样日期)的粪便中检测出SARS-CoV-2阳性。图3),这表明COVID-19患者的一部分肠道中存在SARS-CoV-2。这些受试者都没有胃肠道症状。宏基因组测序未在其他8名患者的粪便中检测到SARS-CoV-2 (在线补充图1).虽然我们的序列深度不允许我们在粪便样本中识别SARS-CoV-2的全长基因组,但有趣的是,我们观察到,在基线时,所有7名患者(宏基因组为SARS-CoV-2阳性)的3 '端区域的覆盖率和密度都高于SARS-CoV-2基因组的5 '端区域(p分别为0.0261和0.0094)。图3 d).中度COVID-19患者12和轻度COVID-19患者15(胸部检查正常)均未表现出胃肠道症状,但在病程的所有时间点,均显示粪便病毒宏基因组中SARS-CoV-2基因组的3 '端覆盖一致(图3A及C).这些发现表明SARS-CoV-2在患者肠道中持续存在。考虑到SARS-CoV-2基因组的3 '端比5 '端覆盖率高是宿主细胞中病毒活跃复制和转录(感染)的一个特征,20.我们的数据强调了COVID-19患者肠道内持续活跃的SARS-CoV-2感染,尽管没有胃肠道症状和轻度COVID-19。相比之下,患者3和7分别在SARS-CoV-2鼻咽清除后1天和10天失去了这种活性病毒感染的特征(图3 b),表明COVID-19患者在病程中消除肠道和呼吸道SARS-CoV-2感染的时间不一致。

COVID-19患者连续粪便中严重急性呼吸综合征冠状病毒2型(SARS-CoV-2)病毒传染性的描述采用粪便病毒RNA宏基因组覆盖谱研究SARS-CoV-2病毒在肠道中的感染性。(A)在咽拭子检测结果为阴性之前,表现出SARS-CoV-2基因组3 ' vs 5 '端覆盖率较高(SARS-CoV-2活性传染性的特征)的患者亚群。(B)表现出更高的SARS-CoV-2基因组3 ' vs 5 '端覆盖率(SARS-CoV-2传染性的特征)的患者亚群,但随着住院时间的推移,在咽拭子为SARS-CoV-2阴性之前逐渐失去了这一特征。(C)咽拭子检测结果为阴性后,SARS-CoV-2基因组3 '端覆盖率高于5 '端覆盖率(SARS-CoV-2传染性的特征)的患者亚群。“第0天”的定义是通过RT-PCR检测到咽喉拭子对SARS-CoV-2呈阴性的日期。(D) SARS-CoV-2基因组5 '端和3 '端在COVID-19粪便病毒RNA宏基因组中的覆盖率和密度。对7例SARS-CoV-2粪便检测阳性患者的基线(住院后第一次收集粪便的日期)粪便病毒RNA宏基因组进行绘制并进行比较。覆盖范围定义为SARS-CoV-2基因组给定基因组区域的霰弹枪读取数。密度定义为SARS-CoV-2基因组特定基因组区域中测序位点的频率。
令人惊讶的是,患者4、11和15的粪便病毒RNA宏基因组继续显示出活跃的病毒感染特征(SARS-CoV-2基因组的3 '端覆盖率和密度高于5 '端)(图3 c),时间点为从咽拭子清除SARS-CoV-2后1至6天。在患者11和15中,咽拭子为阴性后,5 '端在粪便病毒宏基因组中的覆盖率和密度与3 '端相似(例如患者11在第1天和第6天,患者15在第2天,图3 c).这些观察结果表明,这些患者在呼吸道清除SARS-CoV-2后,肠道内的活性病毒感染逐渐消失(从情景3转向情景2)。我们的数据表明,尽管呼吸道清除了SARS-CoV-2,但SARS-CoV-2病毒在人体肠道中持续可检测到,并继续表现出传染性特征,但其转录活性和传染性可能会逐渐下降。没有肠道感染的患者和康复患者的粪便中如此长时间存在活性SARS-CoV-2,突出了粪-口传播的可能性。
与SARS-CoV-2粪便病毒活性相关的粪便微生物组特征
然后,我们研究了传染性高的粪便样本和传染性低或无的粪便样本肠道微生物组的差异,以了解covid -2、微生物组和宿主之间的相互作用。我们对15例COVID-19患者的所有时间点进行了粪便微生物组宏基因组测序,并比较了SARS-CoV-2传染性高和SARS-CoV-2传染性低或无的粪便之间的细菌微生物组组成。我们发现,SARS-CoV-2传染性高的粪便样本中细菌种类的丰度更高Collinsella aerofaciens,Collinsella tanakaei,链球菌对象,摩根氏菌属morganii,与低至无SARS-CoV-2传染性样本比较(LefSE分析,LDA效应量>2,FDR p值<0.05,图4一).在这些物种中,c . aerofaciens而且m . morganii与人类机会性感染有关。21日22美国的对象在上呼吸道和口腔中是大量的殖民者。23在传染性高和传染性低或无的粪便中,粪便宏基因组中的宿主(人类)DNA比例没有差异(在线补充图2).这些数据共同表明,SARS-CoV-2主动感染肠道,并可能对宿主构成额外的威胁。

传染性高的SARS-CoV-2粪便和传染性低到无的SARS-CoV-2粪便的细菌种类和功能能力差异通过对15例COVID-19患者所有时间点粪便的LefSE分析,确定了不同的细菌种类(A)和功能(B)。仅绘制LDA效应值>2和fdr校正p值<0.05的种和功能模块。高传染性定义为粪便病毒RNA宏基因组中SARS-CoV-2基因组的3 ' vs 5 '端覆盖率较高。低传染性定义为粪便病毒RNA宏基因组中SARS-CoV-2基因组的3 '和5 '端覆盖相似或没有覆盖。
相比之下,SARS-CoV-2传染性低或无传染性的粪便样本具有较高的丰度Parabacteroides merdae,拟杆菌stercoris,Alistipes onderdonkii而且螺科细菌1_1_57FAA(LefSE分析,LDA效应量>2,FDR p值<0.05,图4一).细菌成员来自Parabacteroides,拟杆菌而且Lachnospiraceae已知短链脂肪酸(特别是丁酸)的生产者,在增强宿主免疫力方面起着至关重要的作用。24 - 27日Alistipes物种是吲哚阳性,参与血清素前体色氨酸代谢和维持肠道免疫稳态。28 29此外,富含b . stercoris与粪便中SARS-CoV-2病毒含量低也有一定的相关性(斯皮尔曼相关系数rho=−0.26,p=0.06)。我们的数据强调了这些肠道内的有益细菌在对抗肠道内SARS-CoV-2感染方面的潜在有益作用。
我们进一步探讨了具有高传染性特征的粪便和具有低传染性或无传染性特征的粪便之间粪便微生物组功能的差异。我们发现SARS-CoV-2传染性高的粪便样本具有更高丰度的核苷酸代谢功能通路(腺苷核苷酸从头生物合成超通路I、嘌呤核糖核酸降解超通路II、鸟苷核苷酸从头生物合成超通路II、腺苷脱氧核糖核苷酸从头生物合成II、鸟苷脱氧核糖核苷酸从头生物合成II、鸟苷脱氧核糖核苷酸从头生物合成II、鸟苷脱氧核糖核苷酸从头生物合成II、碳水化合物代谢(果糖-6-磷酸的糖酵解II)和氨基酸生物合成(l -赖氨酸生物合成II和l -丝氨酸和甘氨酸生物发生的超通路),与低至无SARS-CoV-2传染性的样品进行比较(LefSE分析,LDA效应量>2,FDR p值<0.05,图4 b).肠道微生物功能中核苷酸和氨基酸生物合成的增加表明了细菌细胞大分子积木生产的潜在增强,而乙醇酸活性的增加表明了细菌能量提取的增强,这是免疫病理条件下肠道细菌的所有基本生命活动。这种与SARS-CoV-2传染性相关的微生物组功能改变可能是COVID-19发病机制和病程的基础,尽管因果关系值得进一步研究。
粪便微生物组与SARS-CoV-2粪便病毒活性的纵向变化
然后,我们研究了粪便微生物群的纵向变化与粪便中活性复制SARS-CoV-2病毒的清除之间的关系。我们随访了3号和7号患者,他们的连续粪便显示出SARS-CoV-2粪便传染性阳性至阴性,以及11号、12号和15号患者,他们的连续粪便不断显示出高病毒传染性的特征。总体而言,无论是否存在粪便病毒感染性,所有患者的粪便微生物组组成均有显著差异(图5),表明在COVID-19的疾病过程中,肠道微生物群不稳定。在粪便中SARS-CoV-2的传染性特征变为阴性后,患者3的有益细菌出现了扩张,Parabacteroides distasonis而且拟杆菌均匀化,炎症相关细菌的收缩,瘤胃球菌属gnavus,在粪便微生物群中。然而,7号病人有短暂性增加p . distasonis伴随着细菌病原体的大量繁殖,肺炎克雷伯菌,枸橼酸杆菌属koseri而且双歧杆菌dentium在清除粪便中的传染性SARS-CoV-2病毒后。肠道炎症和医院感染相关细菌,r . gnavus,hathewayi梭状芽胞杆菌而且肠球菌鸟结核,在SARS-CoV-2传染性持续高的患者的粪便中普遍存在(图5).我们的数据表明,尽管COVID-19患者在通过粪便清除SARS-CoV-2传染性后,肠道微生物群失调可能有所改善,但细菌病原体的继发入侵可能在病程中变得至关重要,因此可能有必要进行临床监测。

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
COVID-19患者粪便微生物群的纵向变化与粪便严重急性呼吸综合征冠状病毒2 (SARS-CoV-2)传染性的相关性患者3和7在随访期间连续大便表现为SARS-CoV-2粪便传染性阳性至阴性,患者11、12和15在随访期间连续大便持续表现为典型的高病毒传染性。只有最丰富的20个物种被绘制并显示了相对丰度。
讨论
我们首次描述了COVID-19患者住院时间内肠道SARS-CoV-2转录活性。COVID-19患者肠道中显著的活性病毒传染性,通过宽靶点霰弹枪病毒宏基因组分析,其特征是SARS-CoV-2基因组的3 '端比5 '端覆盖更高,这表明SARS-CoV-2的胃肠道感染是活跃的,但处于静止状态,因为我们的大多数COVID-19患者没有出现胃肠道症状。在部分COVID-19患者中,即使在呼吸道清除SARS-CoV-2后,这种活跃的病毒感染和复制的转录特征仍然存在于肠道中。我们的数据表明,至少在一部分COVID-19患者中,呼吸道感染和SARS-CoV-2胃肠道感染的清除之间似乎存在长达1周(甚至更长)的滞后窗口期。目前尚不清楚恢复期是否可能是病毒传播的风险,但SARS-CoV-2在肠道中的生命周期似乎比预期的要长。我们的初步观察性研究为COVID-19患者肠道中传染性和延长的SARS-CoV-2病毒活性提供了证据,即使在没有胃肠道表现的情况下。
我们之前的研究显示,15例COVID-19患者中有14例(93%)通过RT-PCR检测粪便样本中SARS-CoV-2阳性。9相比之下,其中7人(47%)通过粪便病毒RNA宏基因组学检测出SARS-CoV-2阳性。只有粪便有丰富的>3.2×104通过粪便病毒RNA宏基因组学检测RT-PCR检测的每mL接种量。霰弹枪宏基因组宽靶点测序在检测目标病毒(SARS-CoV-2)方面的敏感性可能低于RT-PCR,但其优点是可以更广泛地检测SARS-CoV-2基因组的不同区域,从而可以描述病毒的转录活性和感染性。尽管粪便RNA病毒宏基因组可能低估了COVID-19患者肠道中存在SARS-CoV-2病毒的比例,但我们发现,即使没有胃肠道症状,COVID-19患者的粪便中也存在强大的SARS-CoV-2传染性转录特征,表明SARS-CoV-2胃肠道感染处于静止但活跃状态。增加宏基因组测序深度和通量将识别出更多活动性胃肠道感染患者,以及更多SARS-CoV-2病毒变异和突变体。检测到的SARS-CoV-2病毒RNA可能是吞下病毒的残留物。然而,在咽拭子SARS-CoV-2呈阴性后6天仍能检测到病毒RNA (图3 c)表明在粪便中检测到SARS-CoV-2不太可能是人为的。总的来说,我们的数据表明人类肠道是肺外感染部位。
机会性细菌病原体,Collinsella aerofaciens而且摩根氏菌属morganii21日22在SARS-CoV-2传染性高的COVID-19患者粪便样本中富集。链球菌对象在上呼吸道和口腔中有大量的殖民者,23也在这些病人的粪便样本中富集。它的存在表明,在COVID-19环境下,肠外微生物通过或传播到肠道。相比之下,短链脂肪酸和色氨酸生产者,24-29p . merdae,b . stercoris,答:onderdonkii而且Lachnospiraceae细菌1_1_57FAA它们在粪便样本中富集,具有低至无传染性的特征。为了支持这一发现,我们最近的研究还表明,大量的答:onderdonkii而且l .细菌与不那么严重的COVID-19有关。9b . stercoris是一种来自拟杆菌门的细菌物种,已知与小鼠模型中ACE2 (SARS-CoV-2的宿主细胞入口点)的结肠表达抑制有关。30.这些数据强调了有益细菌在抗击SARS-CoV-2感染中的潜在有益作用,以及机会致病菌在SARS-CoV-2感染中的潜在有害作用。然而,这些物种是否可以用于诊断目的尚不清楚。
传染性高的粪便具有较高的核苷酸从头生物合成、氨基酸生物合成和糖酵解微生物功能。其中,腺苷和鸟苷的合成途径明显富集。腺苷和鸟苷是参与嘌呤核苷酸代谢的两种重要代谢物。31腺苷在生理上处于低水平,但在缺氧、缺血、炎症或创伤等病理情况下可以迅速升高,这是一种“警报”或危险信号。32鸟苷及其修饰衍生物8-羟基脱氧鸟苷(8-OHdG)是TLR7的内源性配体。33重要的是,8-OHdG是一种众所周知的氧化性DNA损伤标志物,并诱导产生强烈的细胞因子。33此外,传染性高的粪便中l -丝氨酸的细菌生物合成途径增加。l -丝氨酸可以促进致病菌在发炎肠道中的扩张。34粪便微生物群中微生物功能的增加是细菌生存、生长和代谢所必需的生物活性。这种细菌功能增强可能是SARS-CoV-2在肠道内感染的结果,也可能是增强了COVID-19的病程,这仍有待进一步研究。
呼吸道传播仍是新型冠状病毒的主要传播途径,推测新型冠状病毒可能通过粪口途径传播。35 36研究表明,进入肠腔的新型冠状病毒被结肠液灭活,在新冠肺炎患者粪便标本中未检出传染性病毒。1 37尽管出于安全考虑,我们在处理COVID-19患者粪便样本时,由于方法上的限制,我们无法从COVID-19患者粪便中滴定或分离活病毒,但SARS-CoV-2强大的高粪便病毒转录特征表明它在COVID-19患者肠道中具有活跃的传染性。据此,最近的一项研究能够从一名COVID-19患者的粪便标本中分离出传染性SARS-CoV-2病毒,13进一步支持我们的发现。证据尚不足以制定切实可行的措施,以抑制呼吸道样本结果阴性但粪便样本阳性的患者组的病毒传播和感染。有必要进一步研究揭示SARS-CoV-2在胃肠道中的传染性和发病机制,以及对抗包括胃肠道感染在内的全身感染的措施,如调节人类肠道微生物组。
致谢
作者要感谢在中国香港威尔斯亲王医院和基督教联合医院隔离病房工作的所有医护人员。作者感谢Joyce Mak、Apple CM Yeung、Wendy CS Ho、Miu L Chin、Rity Wong和Vickie Li在本研究中的贡献。作者亦要感谢许辉及周先兰慈善基金有限公司、松鹤有限公司及许明先生对本研究的财政支持。
参考文献
脚注
TZ, QL和FZ贡献相同。
贡献者TZ、QL、FZ分别进行实验、数据分析并撰写稿件。YKY, SSB, ZC, FKLC和PC对手稿进行了修改并提出了批评意见。GC-YL和ET招募患者及临床资料。TZ阐述了概念和假设。SCN设计并监督了这项研究。TZ、QL和FZ对这项工作的贡献相当。
资金这项工作得到陈德宏基金会及香港健康及医学研究基金的支持。
相互竞争的利益没有宣布。
患者发表同意书不是必需的。
伦理批准本研究获香港中文大学-新界东集群联合临床研究伦理委员会(参考编号:2020.076)批准。这项研究是根据《赫尔辛基宣言》进行的。
出处和同行评审不是委托;外部同行评审。
数据可用性声明应合理要求提供数据。